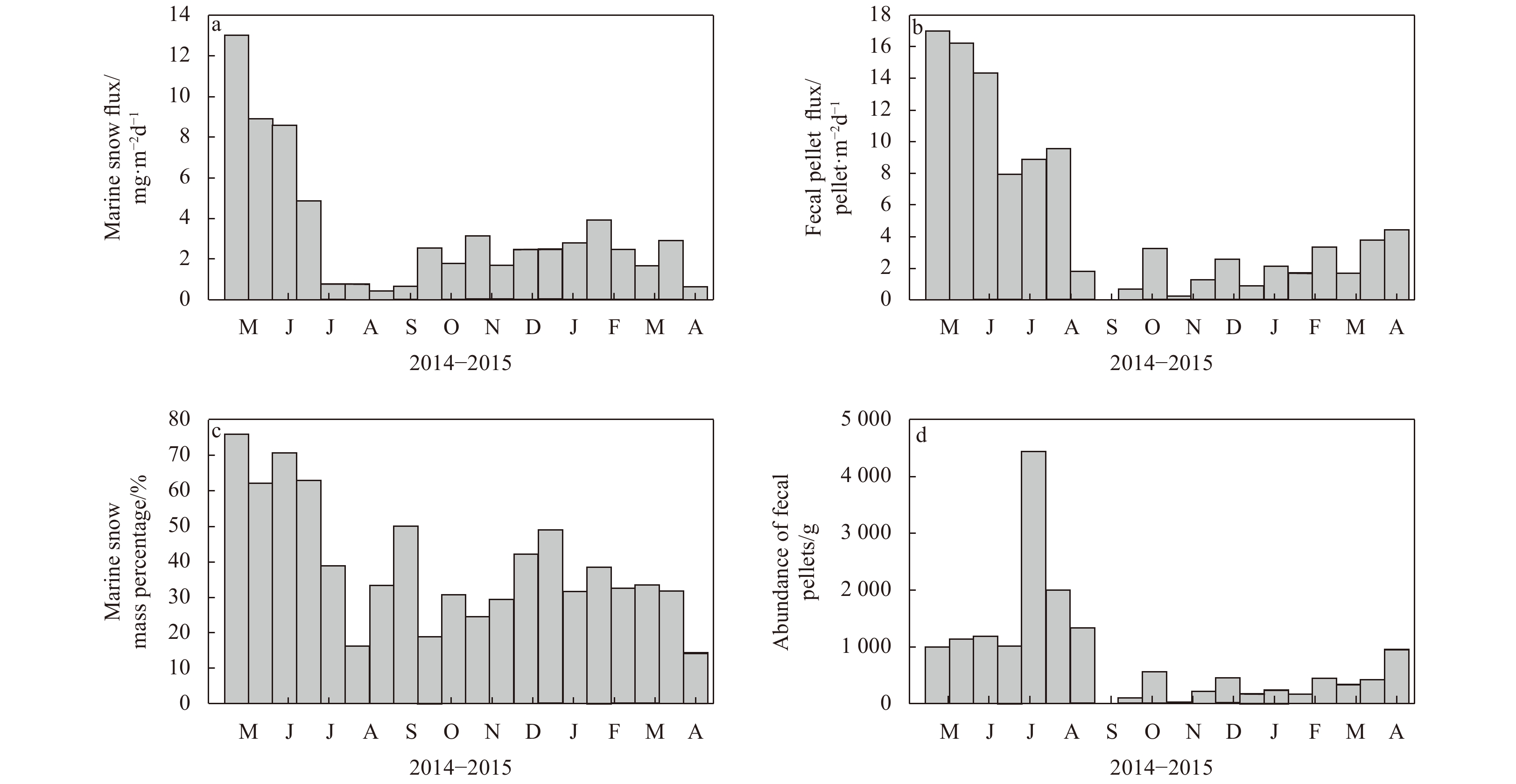
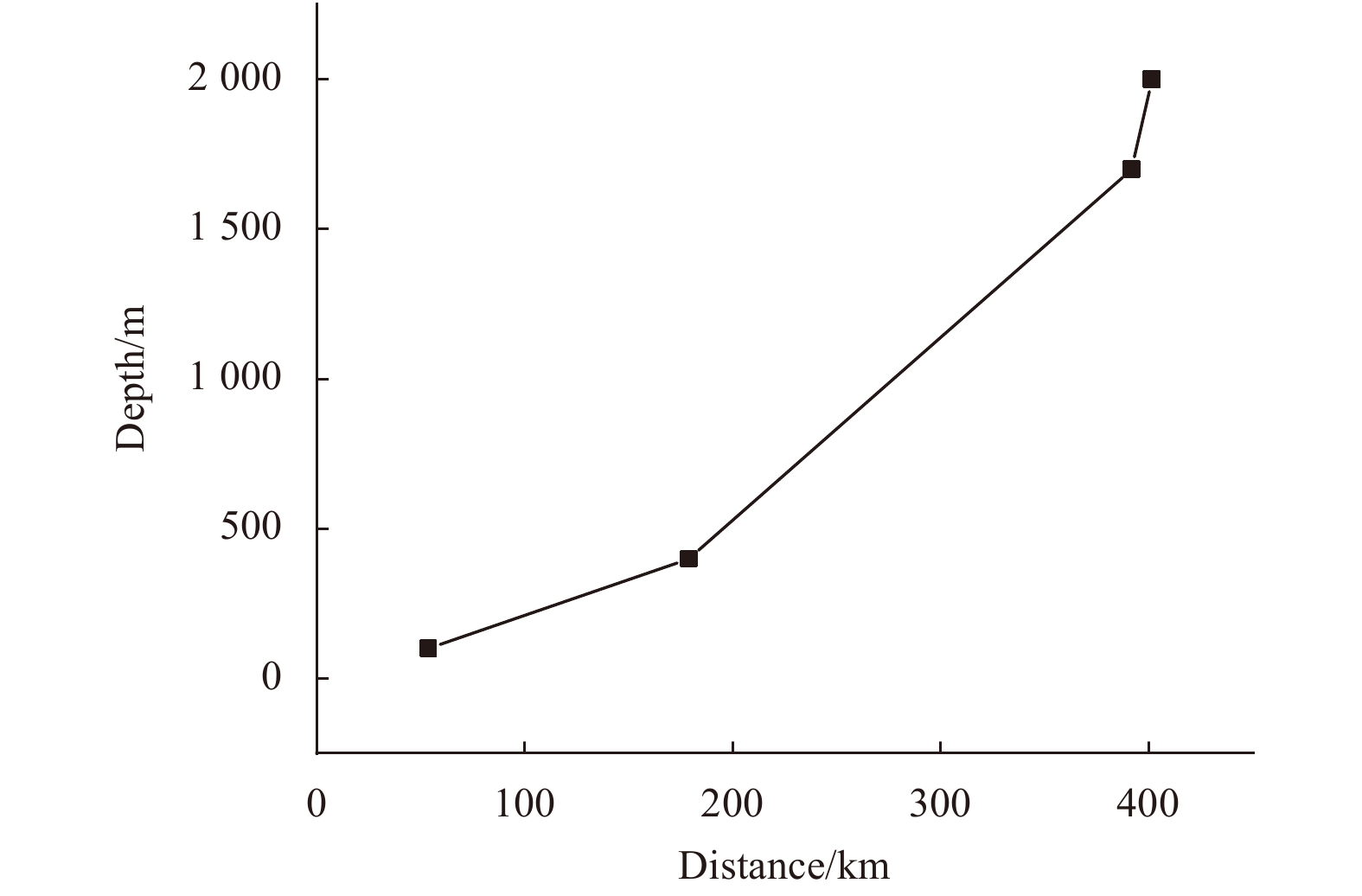
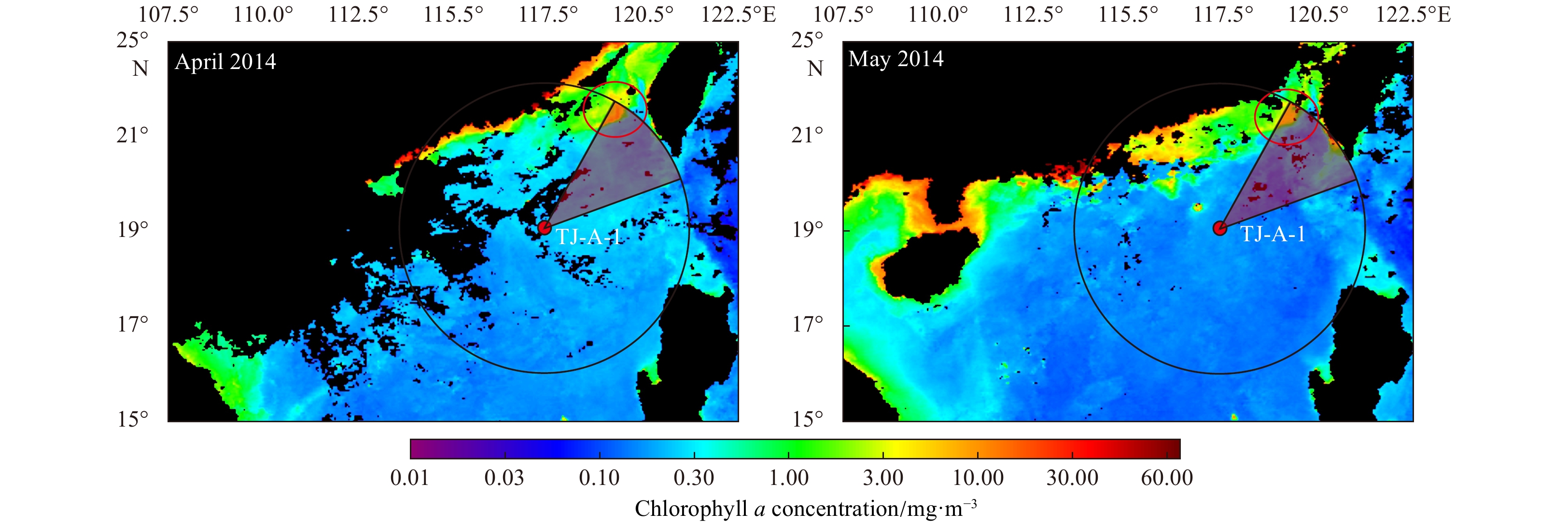
| Citation: | Meng Gao, Baoqi Huang, Zhifei Liu, Yulong Zhao, Yanwei Zhang. Observations of marine snow and fecal pellets in a sediment trap mooring in the northern South China Sea[J]. Acta Oceanologica Sinica, 2020, 39(3): 141-147. doi: 10.1007/s13131-020-1561-9

|

| [1] |
Alldredge A. 1998. The carbon, nitrogen and mass content of marine snow as a function of aggregate size. Deep Sea Research Part I: Oceanographic Research Papers, 45(4–5): 529–541. doi: 10.1016/S0967-0637(97)00048-4
|
| [2] |
Alldredge A L, Silver M W. 1988. Characteristics, dynamics and significance of marine snow. Progress in Oceanography, 20(1): 41–82. doi: 10.1016/0079-6611(88)90053-5
|
| [3] |
Chen Jianfang, Chen Ronghua, Wiesner M G, et al. 2000. Amino acids amino sugars and carbohydrates in settling planktonic tests and their implications for the South China Sea. Oceanologia et Limnologia Sinica (in Chinese), 31(6): 596–603
|
| [4] |
Chen Liumei, Jin Zhangdong. 2013. A review of sediment trap techniques and their applications for understanding of settling particle in lakes and oceans. Journal of Earth Environment (in Chinese), 4(3): 1346–1354
|
| [5] |
Chen Cheng, Mao Zhihua, Han Guoqi, et al. 2016. Latitudinal and interannual variations of the spring phytoplankton bloom peak in the East Asian marginal seas. Acta Oceanologica Sinica, 35(12): 81–88. doi: 10.1007/s13131-016-0867-0
|
| [6] |
Chen Jianfang, Wiesner M G, Li Hongliang, et al. 2015. Biogeochemical fluxes and biological pump composition in the South China Sea. Bulletin of Mineralogy, Petrology and Geochemistry (in Chinese), 34(5): 911–919
|
| [7] |
Chen Jianfang, Zheng Lianfu, Wiesner M G, et al. 1998. Estimations of primary production and export production in the South China Sea based on sediment trap experiments. Chinese Science Bulletin, 43(7): 583–586. doi: 10.1007/BF02883645
|
| [8] |
Chung Y, Chang H C, Hung G W. 2004. Particulate flux and 210Pb determined on the sediment trap and core samples from the northern South China Sea. Continental Shelf Research, 24(6): 673–691. doi: 10.1016/j.csr.2004.01.003
|
| [9] |
Dubischar C D, Bathmann U V. 2002. The occurrence of faecal material in relation to different pelagic systems in the Southern Ocean and its importance for vertical flux. Deep Sea Research Part II Topical Studies in Oceanography, 49(16): 3229–3242. doi: 10.1016/S0967-0645(02)00080-2
|
| [10] |
Ducklow H W, Steinberg D K, Buesseler K O. 2001. Upper ocean carbon export and the biological pump. Oceanography, 14(4): 50–58. doi: 10.5670/oceanog.2001.06
|
| [11] |
Graf G. 1989. Benthic-pelagic coupling in a deep-sea benthic community. Nature, 341(6241): 437–439. doi: 10.1038/341437a0
|
| [12] |
Honjo S, Doherty K W. 1988. Large aperture time-series sediment traps; design objectives, construction and application. Deep Sea Research Part A. Oceanographic Research Papers, 35(1): 133–149. doi: 10.1016/0198-0149(88)90062-3
|
| [13] |
Hung C C, Gong G C. 2010. POC/234Th ratios in particles collected in sediment traps in the northern South China Sea. Estuarine, Coastal and Shelf Science, 88(3): 303–310. doi: 10.1016/j.ecss.2010.04.008
|
| [14] |
Lalande C, Nöthig E M, Bauerfeind E, et al. 2016. Lateral supply and downward export of particulate matter from upper waters to the seafloor in the deep eastern Fram Strait. Deep Sea Research Part I: Oceanographic Research Papers, 114: 78–89. doi: 10.1016/j.dsr.2016.04.014
|
| [15] |
Lin Huiling, Liu J T, Hung Gwowei. 2005. Foraminiferal shells in sediment traps: Implications of biogenic particle transport in the Kao-ping submarine canyon, Taiwan. Continental Shelf Research, 25(18): 2261–2272. doi: 10.1016/j.csr.2005.09.001
|
| [16] |
Liu Jianguo, Clift P D, Yan Wen, et al. 2014. Modern transport and deposition of settling particles in the northern South China Sea: Sediment trap evidence adjacent to Xisha Trough. Deep Sea Research Part I: Oceanographic Research Papers, 93: 145–155. doi: 10.1016/j.dsr.2014.08.005
|
| [17] |
Liu Zhifei, Colin C, Li Xiajing, et al. 2010. Clay mineral distribution in surface sediments of the northeastern South China Sea and surrounding fluvial drainage basins: Source and transport. Marine Geology, 277(1–4): 48–60. doi: 10.1016/j.margeo.2010.08.010
|
| [18] |
Liu Zhifei, Zhao Yulong, Colin C, et al. 2016. Source-to-sink transport processes of fluvial sediments in the South China Sea. Earth-Science Reviews, 153: 238–273. doi: 10.1016/j.earscirev.2015.08.005
|
| [19] |
Longhurst A R, Harrison W G. 1988. Vertical nitrogen flux from the oceanic photic zone by diel migrant zooplankton and nekton. Deep Sea Research Part A. Oceanographic Research Papers, 35(6): 881–889. doi: 10.1016/0198-0149(88)90065-9
|
| [20] |
Matsueda H, Handa N, Inoue I, et al. 1986. Ecological significance of salp fecal pellets collected by sediment traps in the eastern North Pacific. Marine Biology, 91(3): 421–431. doi: 10.1007/BF00428636
|
| [21] |
Miquel J C, Gasser B, Martín J, et al. 2015. Downward particle flux and carbon export in the Beaufort Sea, Arctic Ocean; the role of zooplankton. Biogeosciences, 12(16): 5103–5117. doi: 10.5194/bg-12-5103-2015
|
| [22] |
Qu Tangdong, Girton J B, Whitehead J A. 2006. Deepwater overflow through Luzon Strait. Journal of Geophysical Research: Oceans, 111: C01002
|
| [23] |
Ran Lihua, Chen Jianfang, Wiesner M G, et al. 2015. Variability in the abundance and species composition of diatoms in sinking particles in the northern South China Sea: Results from time-series moored sediment traps. Deep Sea Research Part II: Topical Studies in Oceanography, 122: 15–24. doi: 10.1016/j.dsr2.2015.07.004
|
| [24] |
Rebesco M, Hernández-Molina F J, Van Rooij D, et al. 2014. Contourites and associated sediments controlled by deep-water circulation processes: State-of-the-art and future considerations. Marine Geology, 352: 111–154. doi: 10.1016/j.margeo.2014.03.011
|
| [25] |
Sampei M, Forest A, Sasaki H, et al. 2009. Attenuation of the vertical flux of copepod fecal pellets under Arctic sea ice: evidence for an active detrital food web in winter. Polar Biology, 32(2): 225–232. doi: 10.1007/s00300-008-0523-z
|
| [26] |
Shao Lei, Li Xuejie, Geng Jianhua, et al. 2007. Deep water bottom current deposition in the northern South China Sea. Science in China Series D: Earth Sciences, 50(7): 1060–1066. doi: 10.1007/s11430-007-0015-y
|
| [27] |
Silver M W, Gowing M M. 1991. The "particle" flux: Origins and biological components. Progress in Oceanography, 26(1): 75–113. doi: 10.1016/0079-6611(91)90007-9
|
| [28] |
Simon M, Grossart H P, Schweitzer B, et al. 2002. Microbial ecology of organic aggregates in aquatic ecosystems. Aquatic Microbial Ecology, 28(2): 175–211
|
| [29] |
Sun Jun, Li Xiaoqian, Chen Jianfang, et al. 2016. Progress in oceanic biological pump. Haiyang Xuebao (in Chinese), 38(4): 1–21
|
| [30] |
Tiselius P, Kuylenstierna M. 1996. Growth and decline of a diatom spring bloom phytoplankton species composition, formation of marine snow and the role of heterotrophic dinoflagellates. Journal of Plankton Research, 18(2): 133–155. doi: 10.1093/plankt/18.2.133
|
| [31] |
Turner J T. 2002. Zooplankton fecal pellets, marine snow and sinking phytoplankton blooms. Aquatic Microbial Ecology, 27: 57–102. doi: 10.3354/ame027057
|
| [32] |
Turner J T. 2015. Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Progress in Oceanography, 130: 205–248. doi: 10.1016/j.pocean.2014.08.005
|
| [33] |
Valdes J R, Price J F. 2000. A neutrally buoyant, upper ocean sediment trap. Journal of Atmospheric and Oceanic Technology, 17(1): 62–68. doi: 10.1175/1520-0426(2000)017<0062:ANBUOS>2.0.CO;2
|
| [34] |
Wan Sui, Jian Zhimin, Cheng Xinrong, et al. 2010. Seasonal variations in planktonic foraminiferal flux and the chemical properties of their shells in the southern South China Sea. Science China Earth Sciences, 53(8): 1176–1187. doi: 10.1007/s11430-010-4039-3
|
| [35] |
Waniek J, Koeve W, Prien R D. 2000. Trajectories of sinking particles and the catchment areas above sediment traps in the northeast Atlantic. Journal of Marine Research, 58(6): 983–1006. doi: 10.1357/002224000763485773
|
| [36] |
Wilson S E, Ruhl H A, Smith K L Jr. 2013. Zooplankton fecal pellet flux in the abyssal northeast Pacific: A 15 year time-series study. Limnology and Oceanography, 58(3): 881–892. doi: 10.4319/lo.2013.58.3.0881
|
| [37] |
Xiang Rong, Liu Jianguo, Wang Dongxiao, et al. 2015. Seasonal flux variability of planktonic foraminifera during 2009-2011 in a sediment trap from Xisha Trough, South China Sea. Aquatic Ecosystem Health & Management, 18(4): 403–413
|
| [38] |
Yang Qingxuan, Liang Xinfeng, Tian Jiwei, et al. 2008. Observation on the current in the northern South China Sea and spectrum analysis. Oceanologia et Limnologia Sinica (in Chinese), 39(6): 561–566
|
| [39] |
Yoon W D, Kim S K, Han K N. 2001. Morphology and sinking velocities of fecal pellets of copepod, molluscan, euphausiid, and salp taxa in the northeastern tropical Atlantic. Marine Biology, 139(5): 923–928. doi: 10.1007/s002270100630
|
| [40] |
Zhang Yanwei, Liu Zhifei, Zhao Yulong, et al. 2014. Mesoscale eddies transport deep-sea sediments. Scientific Reports, 4(1): 5937
|
| [41] |
Zhang Yanwei, Liu Zhifei, Zhao Yulong, et al. 2018. Long-term in situ observations on typhoon-triggered turbidity currents in the deep sea. Geology, 46(8): 675–678. doi: 10.1130/G45178.1
|
| [42] |
Zhao Yulong, Liu Zhifei, Zhang Yanwei, et al. 2015. In situ observation of contour currents in the northern South China Sea: Applications for deepwater sediment transport. Earth and Planetary Science Letters, 430: 477–485. doi: 10.1016/j.jpgl.2015.09.008
|
| [43] |
Zheng Hongbo, Yan Pin. 2012. Deep-water bottom current research in the Northern South China Sea. Marine Georesources & Geotechnology, 30(2): 122–129
|

Figures(6) / Tables(1)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: