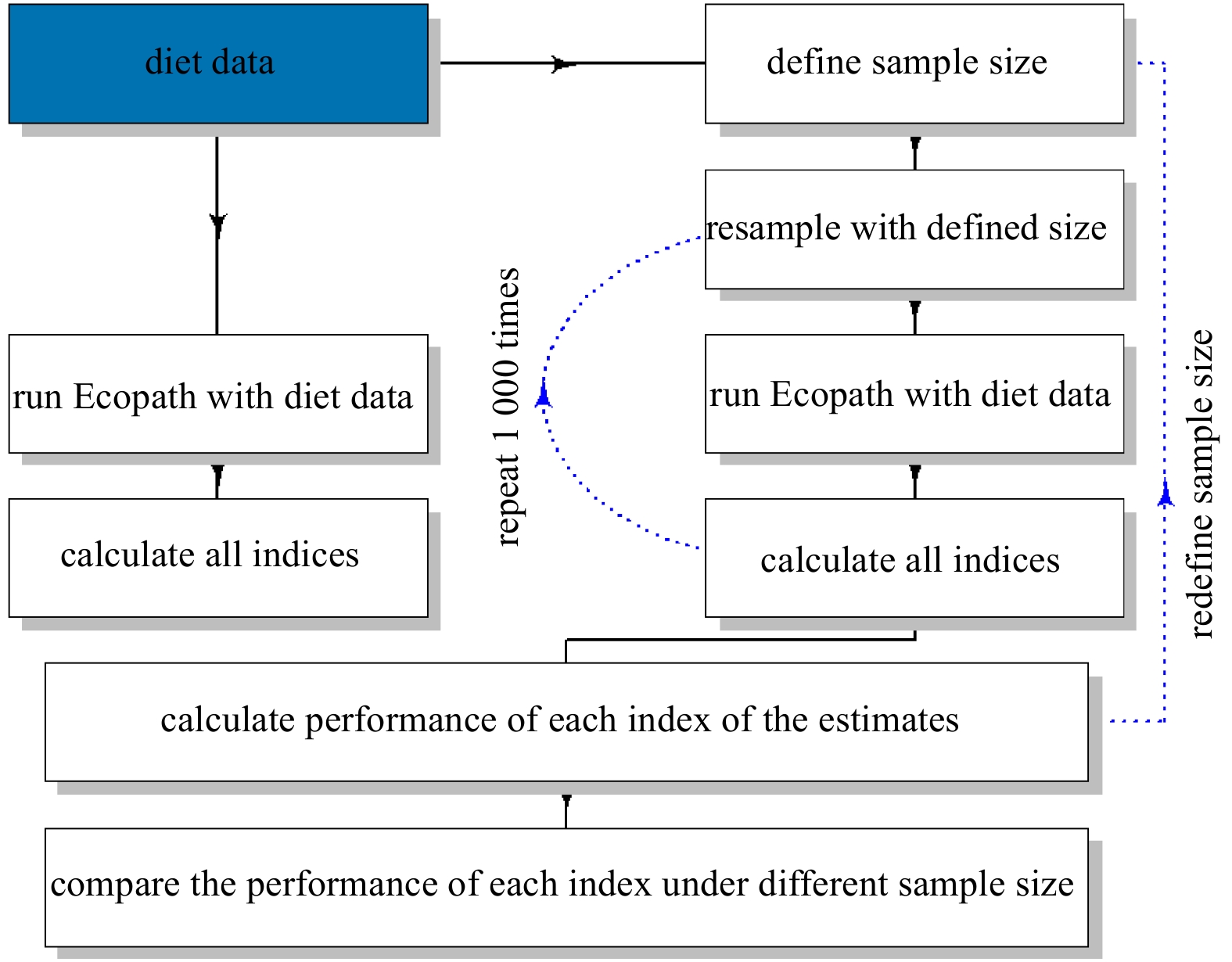
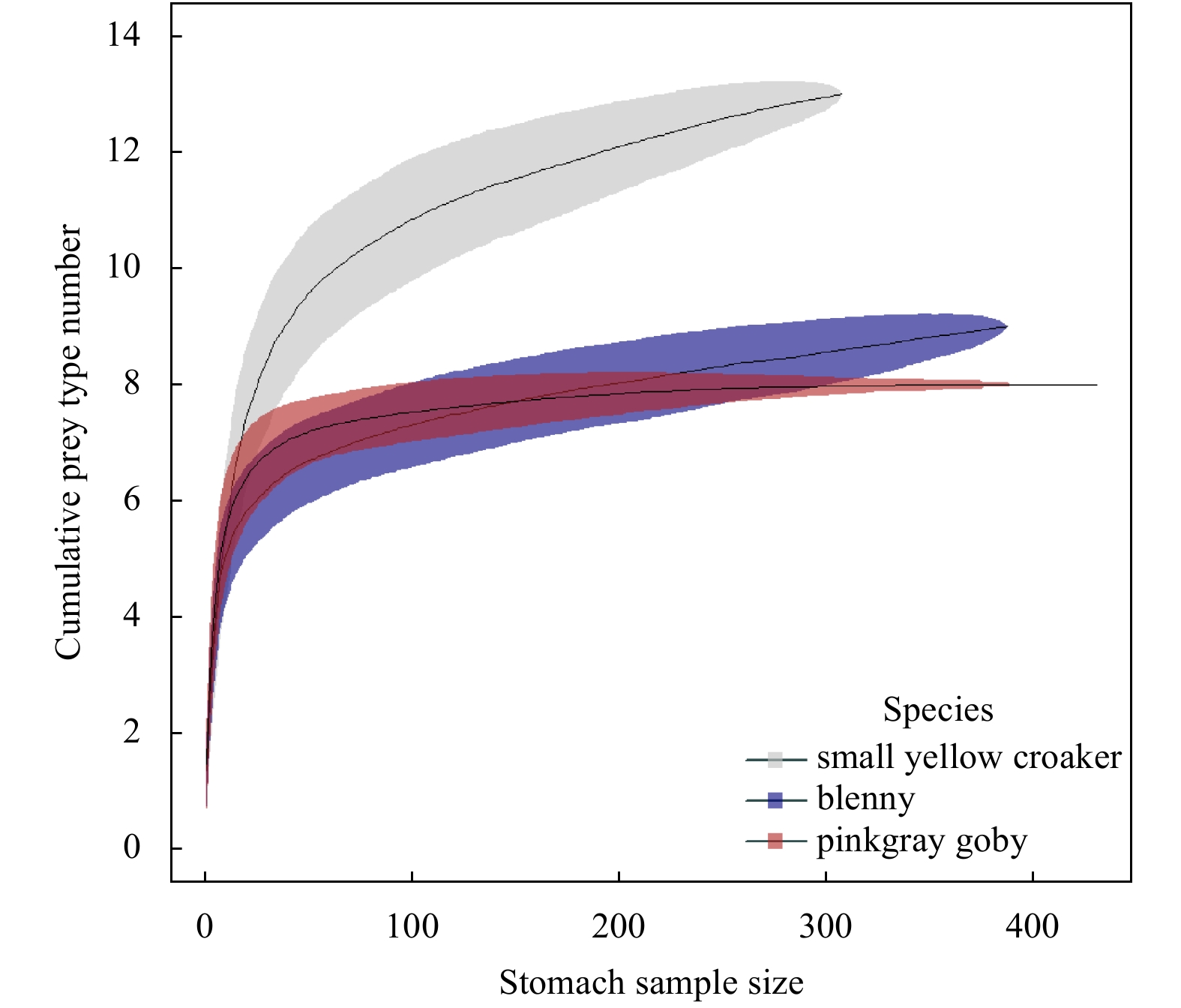
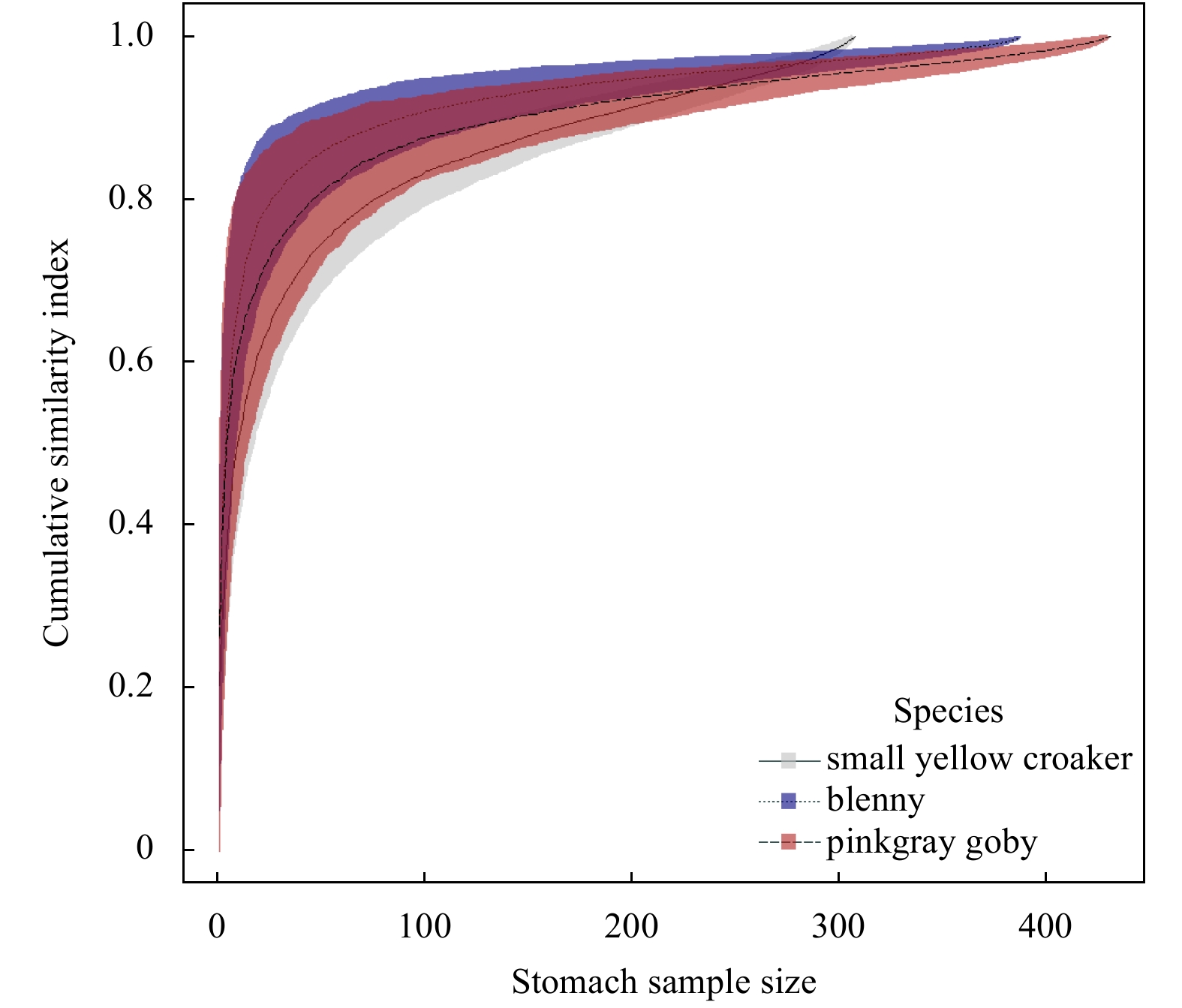
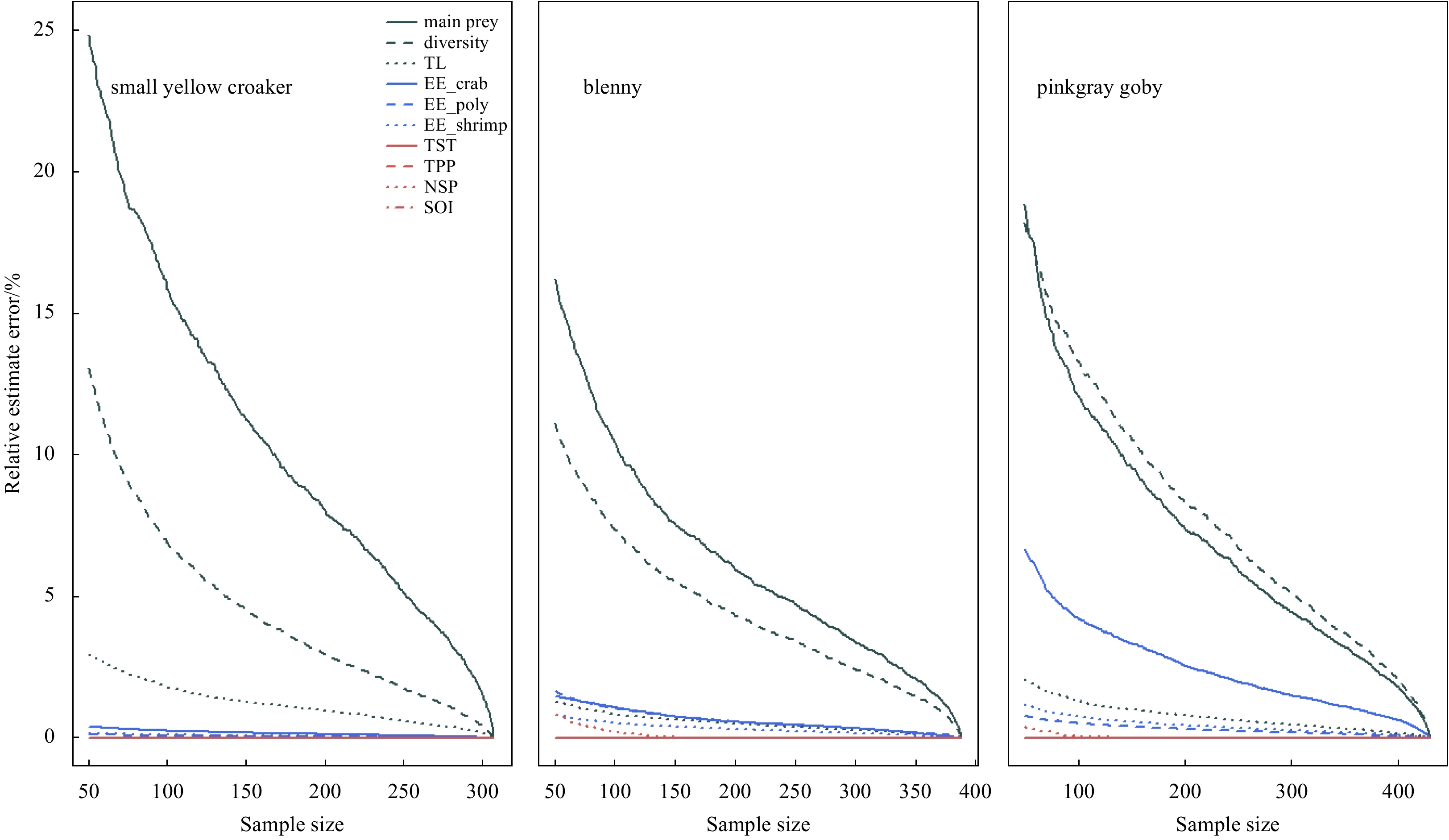
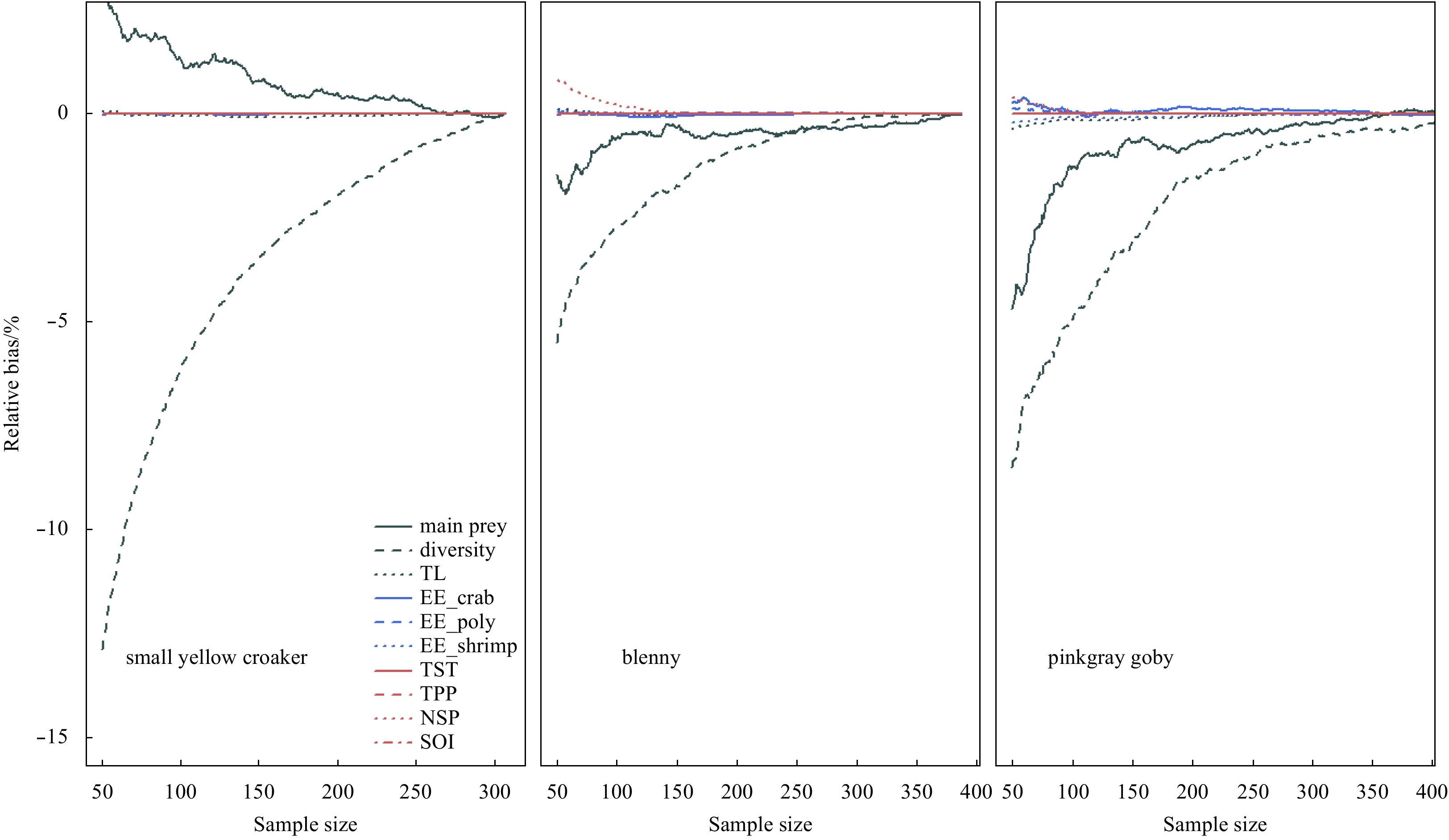
| Citation: | Dongyan Han, Chongliang Zhang, Ying Xue, Binduo Xu, Yiping Ren, Yong Chen. Impacts of sample size for stomach content analysis on the estimation of ecosystem indices[J]. Acta Oceanologica Sinica, 2020, 39(8): 53-61. doi: 10.1007/s13131-020-1633-x

|

| [1] |
Ahlbeck I, Hansson S, Hjerne O. 2012. Evaluating fish diet analysis methods by individual-based modelling. Canadian Journal of Fisheries and Aquatic Sciences, 69(7): 1184–1201. doi: 10.1139/f2012-051
|
| [2] |
Ainsworth C H, Kaplan I C, Levin P S, et al. 2010. A statistical approach for estimating fish diet compositions from multiple data sources: Gulf of California Case study. Ecological Applications, 20(8): 2188–2202. doi: 10.1890/09-0611.1
|
| [3] |
Baker R, Buckland A, Sheaves M. 2014. Fish gut content analysis: robust measures of diet composition. Fish and Fisheries, 15(1): 170–177. doi: 10.1111/faf.12026
|
| [4] |
Bock C, Wermter F C, Mintenbeck K. 2017. MRI and MRS on preserved samples as a tool in fish ecology. Magnetic Resonance Imaging, 38: 39–46. doi: 10.1016/j.mri.2016.12.017
|
| [5] |
Buckland A, Baker R, Loneragan N, et al. 2017. Standardising fish stomach content analysis: the importance of prey condition. Fisheries Research, 196: 126–140. doi: 10.1016/j.fishres.2017.08.003
|
| [6] |
Christensen V, Walters C J. 2004. Ecopath with Ecosim: methods, capabilities and limitations. Ecological Modelling, 172(2–4): 109–139. doi: 10.1016/j.ecolmodel.2003.09.003
|
| [7] |
Crain C M, Kroeker K, Halpern B S. 2008. Interactive and cumulative effects of multiple human stressors in marine systems. Ecology Letters, 11(12): 1304–1315. doi: 10.1111/j.1461-0248.2008.01253.x
|
| [8] |
Cremona F, Järvalt A, Bhele U, et al. 2018. Relationships between fisheries, foodweb structure, and detrital pathway in a large shallow lake. Hydrobiologia, 820(1): 145–163. doi: 10.1007/s10750-018-3648-2
|
| [9] |
Davenport S R, Bax N J. 2002. A trophic study of a marine ecosystem off southeastern Australia using stable isotopes of carbon and nitrogen. Canadian Journal of Fisheries and Aquatic Sciences, 59(3): 514–530. doi: 10.1139/f02-031
|
| [10] |
Elliott J M. 1967. The food of trout (Salmo trutta) in a Dartmoor stream. Journal of Applied Ecology, 4(1): 59–71. doi: 10.2307/2401409
|
| [11] |
Essington T E. 2007. Evaluating the sensitivity of a trophic mass-balance model (Ecopath) to imprecise data inputs. Canadian Journal of Fisheries and Aquatic Sciences, 64(4): 628–637. doi: 10.1139/f07-042
|
| [12] |
Ferry L A, Cailliet G M. 1996. Sample size and data analysis: are we characterizing and comparing diet properly? In: Mackinlay D, Shearer K, eds. Feeding Ecology and Nutrition in Fish, Symposium Proceedings. San Francisco: American Fisheries Society, 71–80
|
| [13] |
Fu Caihong, Travers-Trolet M, Velez L, et al. 2018. Risky business: the combined effects of fishing and changes in primary productivity on fish communities. Ecological Modelling, 368: 265–276. doi: 10.1016/j.ecolmodel.2017.12.003
|
| [14] |
Fulton E A, Smith A D M, Johnson C R. 2004. Biogeochemical marine ecosystem models I: IGBEM—a model of marine bay ecosystems. Ecological Modelling, 174(3): 267–307. doi: 10.1016/j.ecolmodel.2003.09.027
|
| [15] |
Gerringer M E, Popp B N, Linley T D, et al. 2017. Comparative feeding ecology of abyssal and hadal fishes through stomach content and amino acid isotope analysis. Deep Sea Research Part I: Oceanographic Research Papers, 121: 110–120. doi: 10.1016/j.dsr.2017.01.003
|
| [16] |
González J, Ortiz M, Rodríguez-Zaragoza F, et al. 2016. Assessment of long-term changes of ecosystem indexes in Tongoy Bay (SE Pacific coast): based on trophic network analysis. Ecological Indicators, 69: 390–399. doi: 10.1016/j.ecolind.2016.04.019
|
| [17] |
Guesnet V, Lassalle G, Chaalali A, et al. 2015. Incorporating food-web parameter uncertainty into Ecopath-derived ecological network indicators. Ecological Modelling, 313: 29–40. doi: 10.1016/j.ecolmodel.2015.05.036
|
| [18] |
Halouani G, Ben Rais Lasram F, Shin Y J, et al. 2016. Modelling food web structure using an end-to-end approach in the coastal ecosystem of the Gulf of Gabes (Tunisia). Ecological Modelling, 339: 45–57. doi: 10.1016/j.ecolmodel.2016.08.008
|
| [19] |
Han Dongyan, Ma Qiuyun, Xue Ying, et al. 2016. Feeding habits of Amblychaeturichthys hexanema in Jiaozhou Bay based on carbon and nitrogen stable isotope analysis. Periodical of Ocean University of China (in Chinese), 46(3): 67–73
|
| [20] |
Han Dongyan, Xue Ying, Ji Yupeng, et al. 2013. Trophic and spatial niche of five gobiid fishes in Jiaozhou Bay. Journal of Fishery Sciences of China (in Chinese), 20(1): 148–156. doi: 10.3724/SP.J.1118.2013.00148
|
| [21] |
Han Dongyan, Xue Ying, Zhang Chongliang, et al. 2017. A mass balanced model of trophic structure and energy flows of a semi-closed marine ecosystem. Acta Oceanologica Sinica, 36(10): 60–69. doi: 10.1007/s13131-017-1071-6
|
| [22] |
Heymans J J, Coll M, Link J S, et al. 2016. Best practice in Ecopath with Ecosim food-web models for ecosystem-based management. Ecological Modelling, 331: 173–184. doi: 10.1016/j.ecolmodel.2015.12.007
|
| [23] |
Hyslop E J. 1980. Stomach contents analysis—a review of methods and their application. Journal of Fish Biology, 17(4): 411–429. doi: 10.1111/j.1095-8649.1980.tb02775.x
|
| [24] |
Jopp F, Reuter H, Breckling B. 2011. Modelling Complex Ecological Dynamics: an Introduction into Ecological Modelling for Students, Teachers & Scientists. Berlin, Heidelberg: Springer, 1–397
|
| [25] |
Kearney K A. 2017. ecopath_matlab: a matlab-based implementation of the Ecopath food web algorithm. Journal of Open Source Software, 2(9): 1–2. doi: 10.21105/joss.00064
|
| [26] |
Li B, Cao J, Chang J H, et al. 2015. Evaluation of effectiveness of fixed-station sampling for monitoring American lobster settlement. North American Journal of Fisheries Management, 35(5): 942–957. doi: 10.1080/02755947.2015.1074961
|
| [27] |
Liu Ruiyu. 1992. Ecology and Living Resources of Jiaozhou Bay. Beijing: Science Press (in Chinese), 1–429
|
| [28] |
Ma Qiuyun, Jiao Yan, Ren Yiping. 2017. Linear mixed-effects models to describe length-weight relationships for yellow croaker (Larimichthys Polyactis) along the north coast of China. PLoS One, 12(2): e0171811. doi: 10.1371/journal.pone.0171811
|
| [29] |
Masi M D, Ainsworth C H, Jones D L. 2017. Using a Gulf of Mexico Atlantis model to evaluate ecological indicators for sensitivity to fishing mortality and robustness to observation error. Ecological Indicators, 74: 516–525. doi: 10.1016/j.ecolind.2016.11.008
|
| [30] |
McQueen N, Griffiths M H. 2004. Influence of sample size and sampling frequency on the quantitative dietary descriptions of a predatory fish in the Benguela ecosystem. African Journal of Marine Science, 26(1): 205–217. doi: 10.2989/18142320409504058
|
| [31] |
Ofir E, Heymans J J, Shapiro J, et al. 2017. Predicting the impact of Lake Biomanipulation based on food-web modeling—Lake Kinneret as a case study. Ecological Modelling, 348: 14–24. doi: 10.1016/j.ecolmodel.2016.12.019
|
| [32] |
Paloheimo J E, Chen Y. 1996. Estimating fish mortalities and cohort sizes. Canadian Journal of Fisheries and Aquatic Sciences, 53(7): 1572–1579. doi: 10.1139/f96-077
|
| [33] |
Phillips D L, Inger R, Bearhop S, et al. 2014. Best practices for use of stable isotope mixing models in food-web studies. Canadian Journal of Zoology, 92(10): 823–835. doi: 10.1139/cjz-2014-0127
|
| [34] |
Plagányi É E. 2007. Models for an ecosystem approach to fisheries. Rome: FAO, 1–108
|
| [35] |
Raoux A, Tecchio S, Pezy J P, et al. 2017. Benthic and fish aggregation inside an offshore wind farm: which effects on the trophic web functioning?. Ecological Indicators, 72: 33–46. doi: 10.1016/j.ecolind.2016.07.037
|
| [36] |
Shin Y J, Cury P. 2001. Exploring fish community dynamics through size-dependent trophic interactions using a spatialized individual-based model. Aquatic Living Resources, 14(2): 65–80. doi: 10.1016/S0990-7440(01)01106-8
|
| [37] |
Simpson T W, Lin D K J, Chen W. 2001. Sampling strategies for computer experiments: design and analysis. International Journal of Reliability and Applications, 2: 209–240
|
| [38] |
Sokal R R, Rohlf F J. 2012. Biometry: the Principles and Practice of Statistics in Biological Research. 4th ed. New York: W.H.Freeman, 1–102
|
| [39] |
Stergiou K I, Karpouzi V S. 2002. Feeding habits and trophic levels of Mediterranean fish. Reviews in Fish Biology and Fisheries, 11(3): 217–254
|
| [40] |
Wilhm J L. 1968. Use of biomass units in Shannon’s formula. Ecology, 49(1): 153–156. doi: 10.2307/1933573
|
| [41] |
Won N I, Kawamura T, Takami H, et al. 2010. Ontogenetic changes in the feeding habits of the abalone Haliotis discus hannai: field verification by stable isotope analyses. Canadian Journal of Fisheries and Aquatic Sciences, 67(2): 347–356. doi: 10.1139/F09-187
|
| [42] |
Xu Binduo, Zeng Huihui, Xue Ying, et al. 2013. Community structure and species diversity of fish assemblage in the coastal waters of Jiaozhou Bay. Acta Ecologica Sinica (in Chinese), 33(10): 3074–3082. doi: 10.5846/stxb201203040292
|
| [43] |
Xu Binduo, Zhang Chongliang, Xue Ying, et al. 2015. Optimization of sampling effort for a fishery-independent survey with multiple goals. Environmental Monitoring and Assessment, 187(5): 252. doi: 10.1007/s10661-015-4483-9
|
| [44] |
Xue Ying, Jin Xianshi, Zhang Bo, et al. 2004. Ontogenetic and diel variation in feeding habits of small yellow croaker Pseudosciaena polyactis Bleeker in the central part of Yellow Sea. Journal of Fishery Sciences of China (in Chinese), 11(5): 420–425
|
| [45] |
Yuan Yuan, Song Dehai, Wu Wen, et al. 2016. The impact of anthropogenic activities on marine environment in Jiaozhou Bay, Qingdao, China: a review and a case study. Regional Studies in Marine Science, 8: 287–296. doi: 10.1016/j.rsma.2016.01.004
|
| [46] |
Zeng Xiaoqi, Piao Chenghua, Jiang Wei, et al. 2004. Biodiversity investigation in Jiaozhou bay and neighbouring waters. Periodical of Ocean University of China (in Chinese), 34(6): 977–982
|
| [47] |
Zeng Huihui, Xu Binduo, Xue Ying, et al. 2012. Study on fish species composition and seasonal variation in the shallow waters of Jiaozhou bay. Periodical of Ocean University of China (in Chinese), 42(1): 67–74
|
Figures(8) / Tables(2)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: