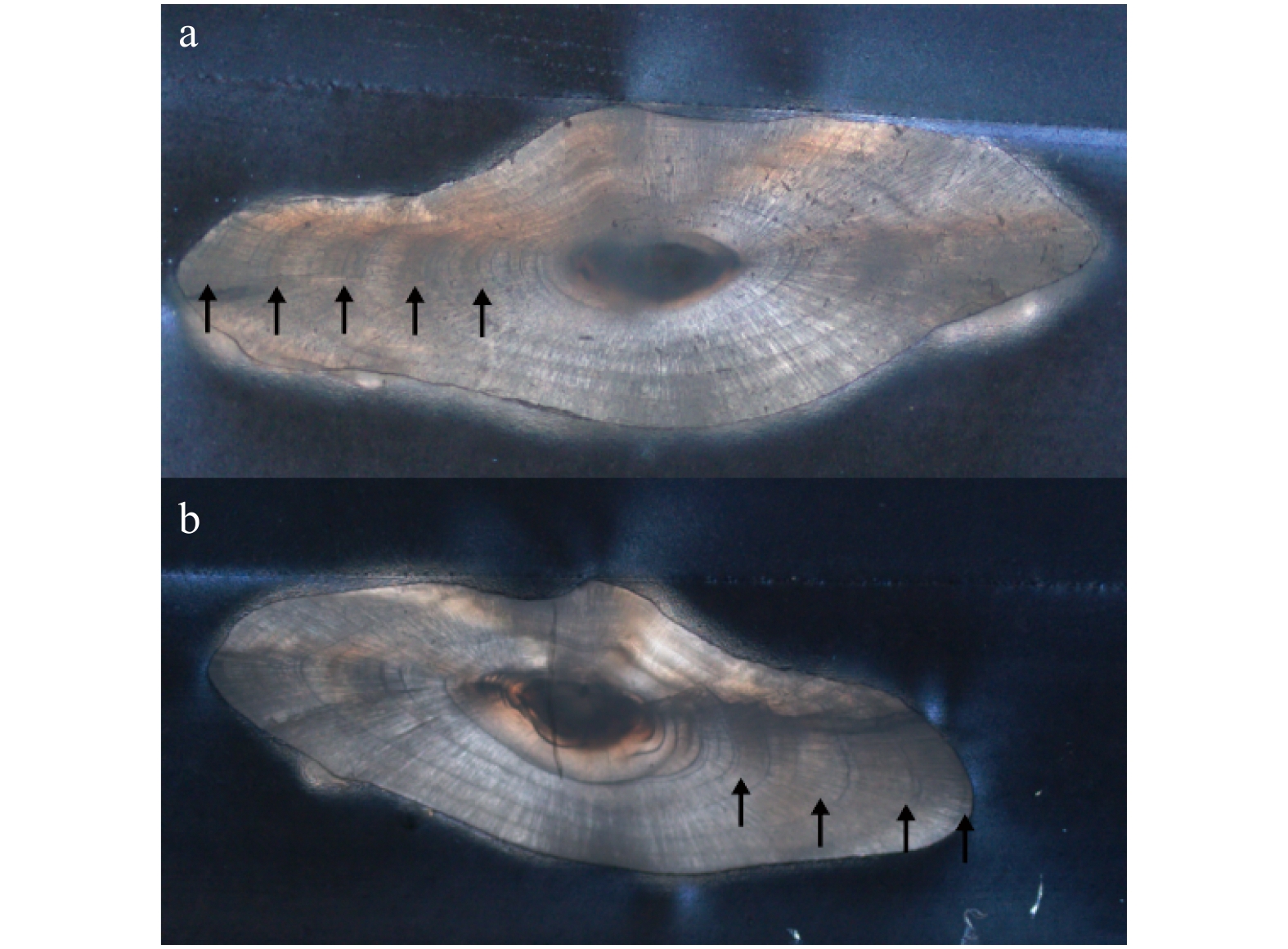
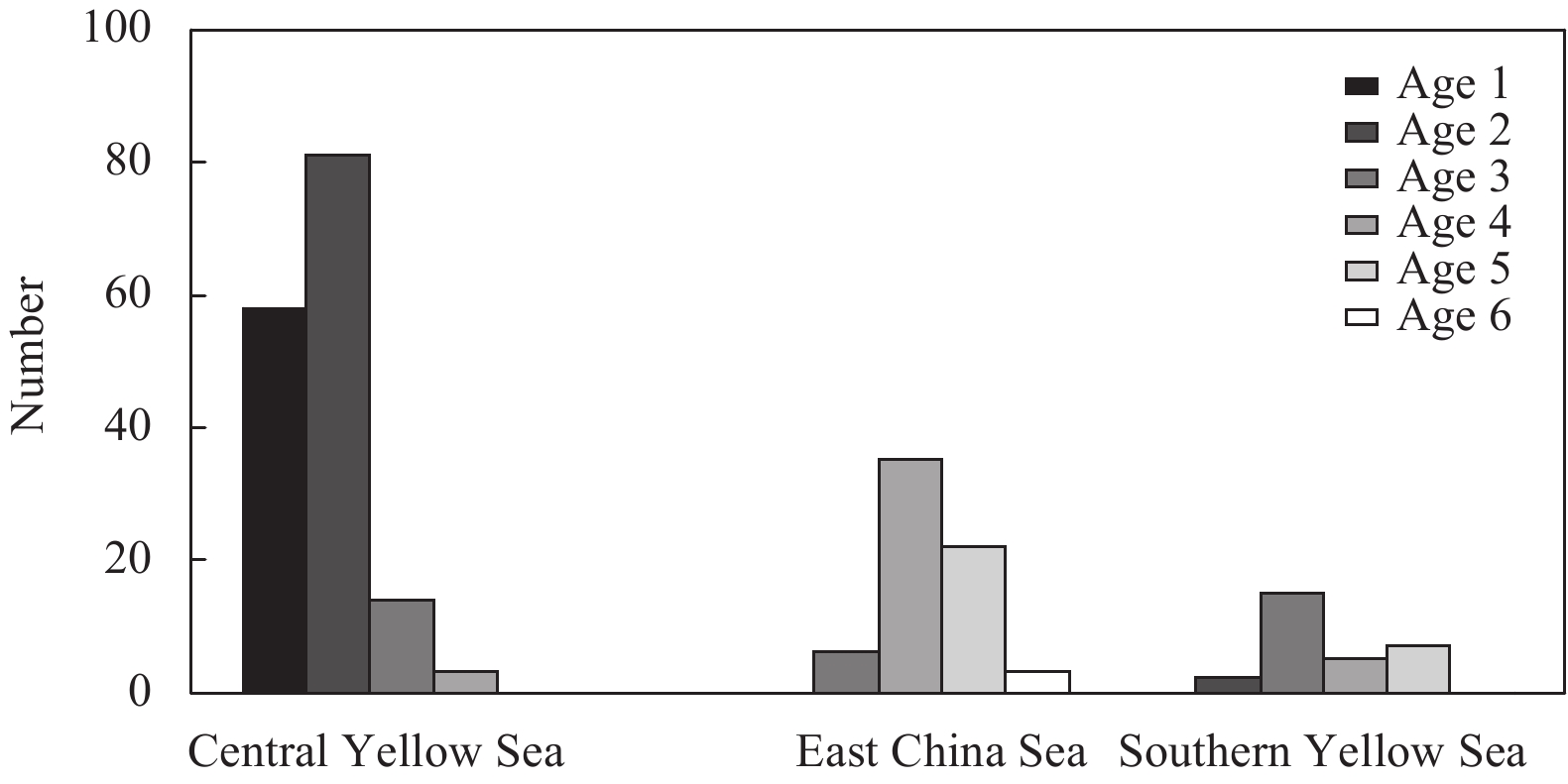
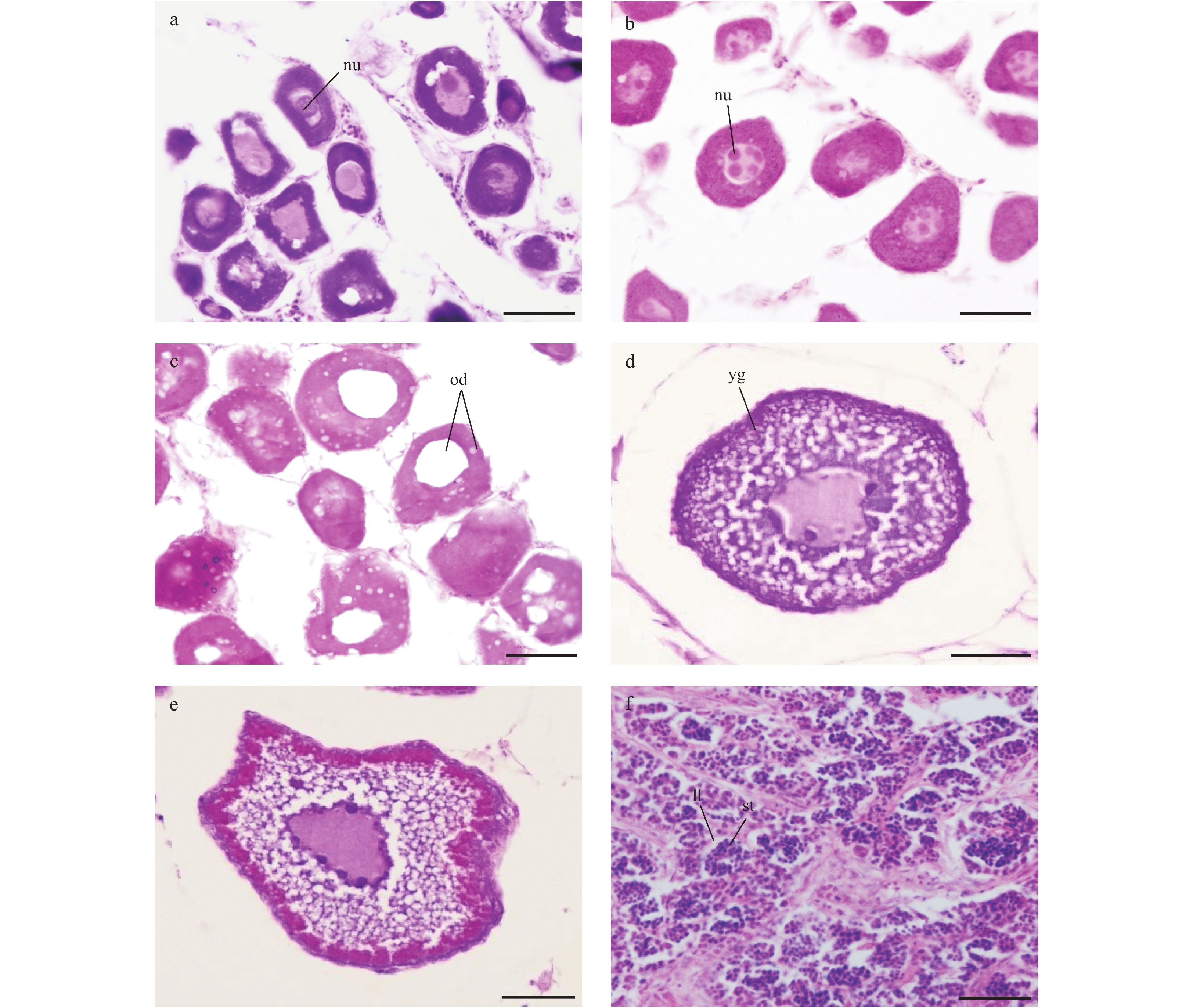
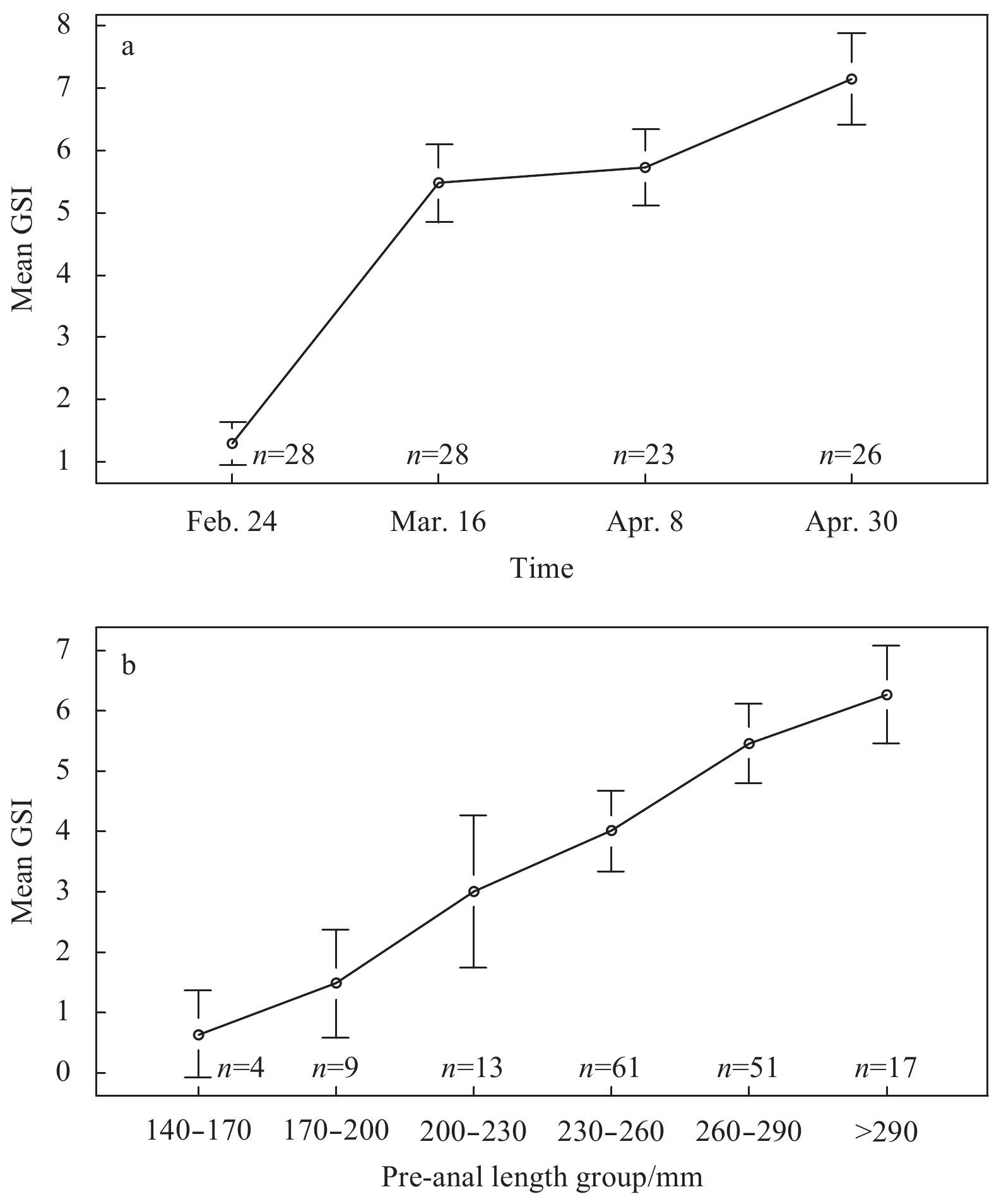
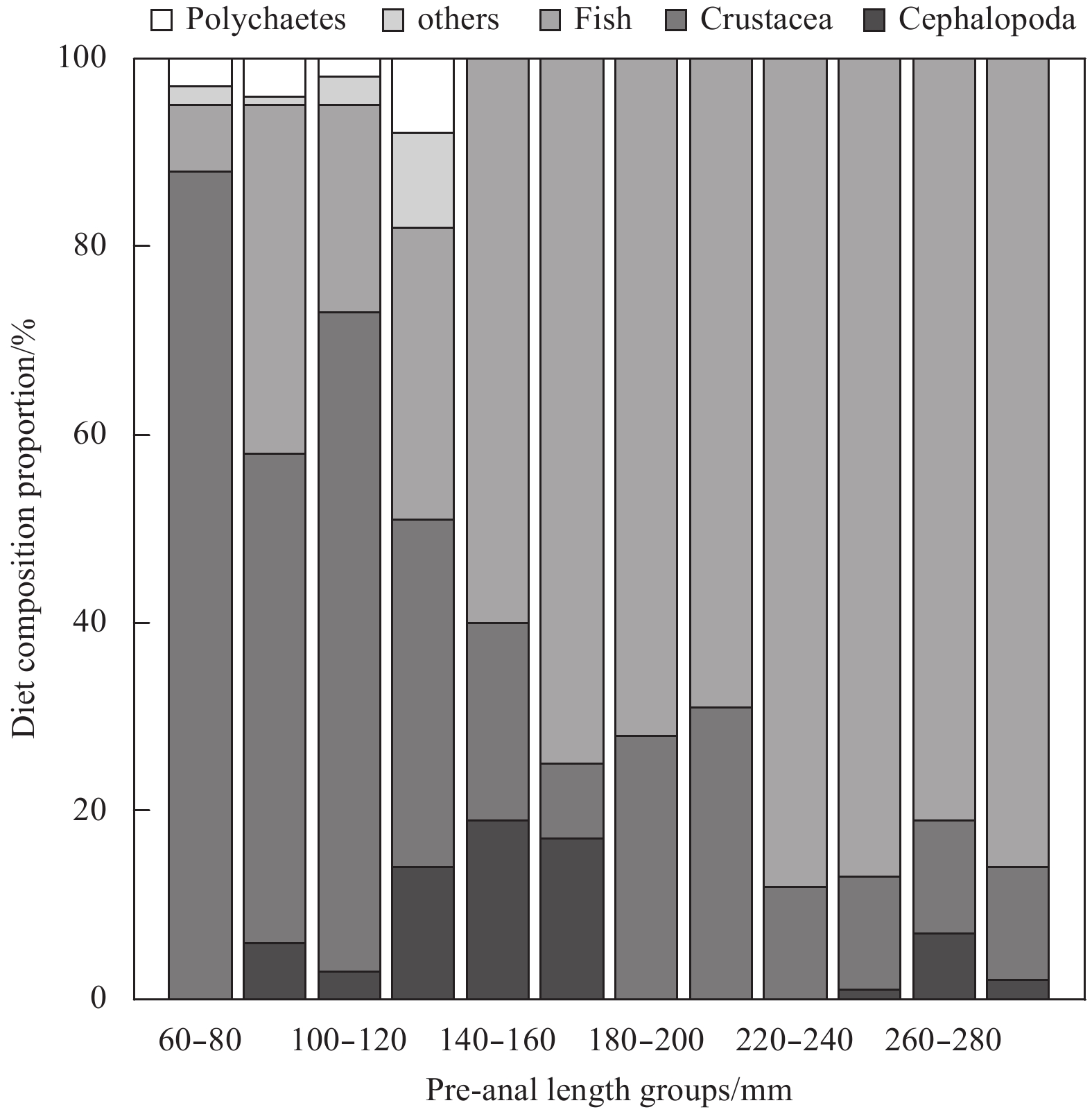
| Citation: | Xiuxia MU, Chi ZHANG, Ying XUE, Chongliang ZHANG, Binduo XU, Yiping REN. Fishery biology of whitespotted conger Conger myriaster (Brevoort, 1856) in the Yellow Sea and East China Sea[J]. Acta Oceanologica Sinica, 2019, 38(6): 18-24. doi: 10.1007/s13131-018-1271-8

|
|
Bae J H, Bae H J, Park H M, et al. 2018. Age determination and growth estimates of the white-spotted conger eel, Conger myriaster (Brevoort, 1856) in marine waters of South Korea. Journal of Applied Ichthyology, 34(3): 542–549
|
|
Chen Dagang. 1991. Fish Ecology in the Yellow Sea and Bohai Sea (in Chinese). Beijing: China Ocean Press, 218–221
|
|
Choi J H, Choi S H, Kim J B, et al. 2008. Feeding ecology of the white-spotted conger eel (Conger myriaster) in the Southern Sea of Korea. Korean Journal of Fisheries and Aquatic Sciences (in Korean), 41(4): 282–288
|
|
Correia A T, Manso S, Coimbra J. 2009. Age, growth and reproductive biology of the European conger eel (Conger conger) from the Atlantic Iberian waters. Fisheries Research, 99(3): 196–202
|
|
Gorie S, Nagasawa K. 2010. Habitat preference and feeding habits of juvenile whitespotted conger Conger myriaster in the western Seto Inland Sea, Japan. The Aquaculture (in Japanese), 58(2): 167–179
|
|
Gorie S, Tanda M, Katayama S. 2004. Age and growth of white-spotted conger Conger myriaster collected in northeast Harima Nada, Seto Inland Sea. Aquaculture Science (in Japanese), 52(4): 407–411
|
|
Gorie S, Tanda M, Nagasawa K. 2010. Movement and growth of whitespotted conger Conger myriaster in the western Seto Inland Sea, Japan. Aquaculture Science (in Japanese), 58(2): 233–242
|
|
Harada M, Tokai T, Uchida K, et al. 2006. Distribution of white-spotted conger eel Conger myriaster and hagfish Eptatretus burgeri in the shallow region of Tokyo Bay. Nippon Suisan Gakkaishi (in Japanese), 72(5): 894–904
|
|
Huh S H, Kwak S N. 1998. Feeding habits of Conger myriaster in the eelgrass (Zostera marina) bed in Kwangyang Bay. Korean Journal of Fisheries and Aquatic Sciences (in Korean), 31(5): 665–672
|
|
Katayama S, Ishida T, Goto K, et al. 2002. A new aging technique by UV light observation of burnt otoliths for the conger eel Conger myriaster (Brevoort). Ichthyological Research, 49(1): 81–84
|
|
Katayama S, Ishida T, Shimizu Y, et al. 2004. Seasonal change in distribution of Conger eel Conger myriaster off the Pacific coast south of Tohoku, north-western Japan. Fisheries Science, 70(1): 1–6
|
|
Kawazu M, Kameda T, Kurogi H, et al. 2015. Biological characteristics of Conger myriaster during the initial stage of spawning migration in the East China Sea. Fisheries Science, 81(4): 663–671
|
|
Kim Y H, Lee E H, Kim J N, et al. 2011. Age and growth of whitespotted conger Conger myriaster in the southern Coastal Waters of Korea. Korean Journal of Fisheries and Aquatic Sciences (in Korean), 44(6): 689–694
|
|
Kimura Y, Ishikawa S, Tokai T, et al. 2004. Early life history characteristics and genetic homogeneity of Conger myriaster leptocephali along the east coast of central Japan. Fisheries Research, 70(1): 61–69
|
|
Kurogi H, Mochioka N, Okazaki M, et al. 2012. Discovery of a spawning area of the common Japanese conger Conger myriaster along the Kyushu-Palau Ridge in the western North Pacific. Fisheries Science, 78(3): 525–532
|
|
Liu Jing, Ning Ping. 2011. Species composition and faunal characteristics of fishes in the Yellow Sea. Biodiversity Science (in Chinese), 19(6): 764–769
|
|
Liu Xifang, Liu He, Xue Ying, et al. 2015. Feeding ecology of Conger myriaster in Haizhou Bay. Journal of Fishery Sciences of China (in Chinese), 22(3): 517–527
|
|
Ogle D H. 2016. Introductory Fisheries Analyses with R. Boca Raton: CRC Press
|
|
Okamura A, Utoh T, Zhang Huan, et al. 2000. Seasonal changes in maturity in the conger eel Conger myriaster at the Pacific coast of Atsumi Peninsula, central Japan. Nippon Suisan Gakkaishi (in Japanese), 66(3): 412–416
|
|
Pauly D. 1981. The relationships between gill surface area and growth performance in fish: a generalization of von Bertalanffy’s theory of growth. Meeresforschung, 28(4): 251–282
|
|
Ricker W E. 1975. Computation and interpretation of biology statistics of fish populations. Bulletin of the Fisheries Research Board of Canada, 191: 200
|
|
Tang Yanli, Sun Guowei, Zhao Tongyang, et al. 2010. Mesh size selectivity of Conger myriaster pot. Journal of Fishery Sciences of China (in Chinese), 17(1): 136–142
|
|
Tang Yimin, Wu Changwen. 1988. On biologic habits and resources distribution of common Japanese conger Astroconger myriaster (Brevoort). Journal of Zhejiang College of Fisheries (in Chinese), 7(1): 19–26
|
|
Tokimura M. 2001. Conger-eel fisheries and fisheries resources in the East China Sea. Nippon Suisan Gakkaishi (in Japanese), 67(1): 125–126
|
|
Utoh T, Horie N, Okamura A, et al. 2003. Oogenesis in the common Japanese conger Conger myriaster. Fisheries Science, 69(1): 181–188
|
|
Utoh T, Okamura A, Yamada Y, et al. 2004. Reproductive cycle in reared male common Japanese conger, Conger myriaster. Aquaculture, 240(1–4): 589–605
|
|
Von Bertalanffy L. 1951. Theoretische Biologie: II. Band, Stoffwechsel, Wachstum. 2nd ed. Bern, Switzerland: A Francke AG Verlag, 418
|
|
Yagi Y, Kodono N, Kinoshita I, et al. 2010. Late-stage metamorphosing Conger myriaster leptocephali collected in a river estuary of Ariake Bay, Japan. Ichthyological Research, 57(3): 310–313
|
|
Yamada U. 1986. Conger myriaster. In: Okamura O, ed. Fishes of the East China Sea and the Yellow Sea (in Japanese). Nagasaki: Seikai National Fisheries Research Laboratory, 68–69
|
|
Zhang Bo, Tang Qisheng. 2003. Feeding habits of six species of eels in East China Sea and Yellow Sea. Journal of Fisheries of China (in Chinese), 27(4): 307–314
|
|
Zhang Chunguang. 2010. Fauna Sinica: Osteichthyes, Anguilliformes and Notacanthiformes (in Chinese). Beijing: Science Press, 199–203
|
|
Zhang Shiyi, Xiao Zhenyi, Zhang Youwei. 1977. Leptocephalus of the Aetro conger myriaster from the East China Sea. Acta Zoologica Sinica (in Chinese), 1: 117–118
|

Figures(6) / Tables(4)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: