Key Laboratory of Marine Bioresource Restoration and Habitat Reparation in Liaoning Province, Dalian Ocean University, Dalian 116023, China
2.
Marine and Fisheries Technology Center of Pan Shan Country, Panjin 124000, China
Funds:
The Dalian Science Fund for Distinguished Young Scholars under contract No. 2016RJ06; the Natural Science Foundation of Liaoning Province under contract No. 201602107; the Superior College Science Technology Research Project of Liaoning Province under contract No. ZL201701; the National Natural Science Foundation of China under contract No. 41306138.
Rapid amplification of cDNA ends (RACE) and real-time polymerase chain reaction (RT-PCR) were carried out to analyze the CYP4 gene expression in polychaete Marphysa sanguinea exposed to benzo[a]pyrene (BaP) in this study. The full length of MsCYP4 cDNA was 2 470 bp, and it encoded 512 amino acids. The deduced amino acid sequence showed 47% identity with CYP4F from frog Xenopus tropicalis and shared high homology with other known CYP4 sequences. To analyse the role of CYP4 in protecting M. sanguinea from BaP exposure, three BaP groups were established: 0.5, 5 and 50 μg/L. Polychaetes were sampled after 3, 7 and 12 d. At 0.5 μg/L, the effect of BaP on MsCYP4 gene expression increased with time prolonged. MsCYP4 gene expression curve showed U-shaped trend with time in 5 and 50 μg/L BaP groups. Therefore, MsCYP4 gene may play an important role in maintaining the balance of cellular metabolism and protecting M. sanguinea from BaP toxicity.
Polycyclic aromatic hydrocarbons (PAHs), which are composed of two or more aromatic nuclei, have been regarded as a primary pollution source in marine coastal environment (Martins et al., 2013). The oil spill in the Gulf of Mexico (2010) as well as in the Bohai Bay of China (2011), and ship wrecks such as the Prestige accident in 2002, the Erika accident in 1999 and the Volganeft-139 accident in 2010 have caused several millions of crude oil leaking into the sea, and its main content PAHs made serious impact on the marine environment in those areas. PAHs are persistent, toxic, and contain carcinogenic compounds that can attack DNA by forming PAH-DNA adducts and cause disorders in marine animals (Tsai et al., 2007). These compounds can also be transferred throughout the food chain and threaten higher levels of organisms, through biomagnification; as a result, various disorders, such as decreased fertility, obesity, and abnormal organ development, occured (Kraugerud et al., 2012; Lyche et al., 2011, 2013). Low-molecular-weight PAHs volatilize into the atmosphere, but high-molecular-weight PAHs subside in water and adsorb on the surface of sediment particles; the adsorbed PAHs then accumulate in estuarine sediments and absorb by the benthonic animal (Barakat et al., 2011; Tobiszewski and Namieśnik, 2012).
Cytochrome P450 (CYP) is a superfamily of protoheme oxygenase that catalyze a high range of reactions, such as phase I oxidation, production, and reduction of endogenous substances, including vitamins, steroids, fatty acids, and prostaglandins. These enzymes also play an essential role in metabolizing xenobiotics, such as PAHs, to detoxified forms, which are hydrophilic, or to harmful reactive intermediates (Bach et al., 2005; Scornaienchi et al., 2010). Study showed that CYP1A1 plays a protective role against oral BaP toxicity in aorta ROS production contributed to BaP-exacerbated atherosclerosis, but no CYP1A has been identified in invertebrates (Uno et al., 2014). CYP4 have been proven to be responsible for CYP1A-like responses, and more importantly these enzymes were widely found in bacteria, fungi, plants, vertebrates and invertebrates; hence, CYP4 constitute a diverse and large number of subfamilies, such as CYP4B, CYP4F, CYP4T, CYP4V, CYP4X, and CYP4Z (Kirischian and Wilson, 2012). Meanwhile, a high diversity of CYP4 genes has also been determined in marine invertebrates such as mussels, oysters, shrimp and polychaete (Guo et al., 2013; Zanette et al., 2010). The CYP4 enzymes present a high degree of structural diversity and comprise numerous homologous members among invertebrates. The protective role of CYP4 against BaP has also been explored in other marine invertebrates, including clam Ruditapes philippinarum, polychaete Perinereisnuntia, Perinereis aibuhitensis, mollusk Pernaviridis, and scallop Chlamys farreri, the results showed that CYP4 mRNA expression was significantly induced in different PAHs-polluted environments (Miao et al., 2011; Pan et al., 2011; Zhou et al., 2010a, b ). CYP4 play an important role in metabolism of PAHs, Pan et al. (2011) demonstrated that CYP4 mRNA expression was induced significantly in the digestive gland of the clam Ruditapes philippinarum by exposure to 0.2 μg/L BaP. Chen et al. (2012) proposed that the expression pattern of CYP4 in PAH-exposed polychaete Perinereisaibuhitensis changes with time in a dose-dependent manner which petroleum hydrocarbon and combined toxicity treatments elevated the mRNA level of PaCYP4.
Polychaetes, which possess various types of CYP4, have been used as a toxicological animal model. Jørgensen et al. (2005) revealed two CYP4 gene sequences in Nereis virens. CYP4BB2, CYP423A1 and CYP424A1 have been cloned in Perinereis nuntia (Zheng et al., 2013). Polychaetes are important geographically distributed marine invertebrates in mudflats and estuarine sediments. They exhibit strong tolerance and bioavailability to many persistent pollutants (Lewis and Watson, 2012). PAHs can accumulate in their bodies after a long-term exposure. Thus, the polychaete is often considered an indicator organism in the assessments of coastal sediment pollution and model species in toxicological studies (Alam et al., 2010; Dean, 2008; Musale and Desai, 2011). The polychaete Marphysa sanguinea is an important commercial species widely distributed along the coasts of Southeast Asia (Zhao et al., 2016). This species is an essential carrier for the transfer of contaminated sediments to higher trophic levels and key point to detoxifying PAHs in the food chain of marine ecosystems (Onozato et al., 2012). The relationship between organic chemicals, such as PAHs, and the responses to the stress of CYP4 gene expression in M. sanguinea should be analyzed to monitor PAHs pollution and to establish a theoretical basis for understanding the regulation of the body’s detoxification signal transduction system. As PAHs concentrations in aquatic environmental increased because of oil spill, the effects on benthic invertebrates should be experimented, especially on polychaete (Feng et al., 2009; Hong et al., 2012; Jernelöv, 2010; Sim et al., 2010). In our study, the full-length cDNA of CYP4 from M. sanguinea was cloned, and the expression of MsCYP4 mRNA under BaP exposure was examined. This study aimed to elaborate the relationship between the varying levels of CYP4 gene expression in M. sanguinea and PAHs pollution concentration. Our study could also provide a basis for the investigation of stress-related responses, ecological assessment of petroleum pollution, and identification of potential biomarkers of organic pollutants in polychaete.
2.
Materials and methods
2.1
Animals and exposure experiment
Segment worms M. sanguinea (1.5–2.5 g wet weight) were collected in tidal sand flat sediments from Dalian Heishijia coast (39°N, 121°E), water salinity was 32, pH 8.25±0.10, temperature (14±0.5)°C. The animals were transferred from the field to the laboratory with sediments and acclimated in seawater and mud for a week. The seawater was changed daily and kept at a temperature of (14±0.5)°C.
According to People’s Republic of China sea water quality standard and our previous study (Song et al., 2011), three BaP (Sigma, America) concentrations were used: 0.5, 5 and 50 μg/L. A solvent control group (100 μL/L acetone) and a blank group were also set. A total of 0.05 g BaP powder was dissolved in 100 mL acetone to prepare 0.5 g/L stock solution. And then stock solution was added to 500 mL seawater to forming final concentration. Five polychaetes were placed into each plastic beaker. Three replicates were used for each treatment. The seawater was changed daily during the experiment and experimental conditions (salinity, pH, temperature, and density) were the same as those used for acclimation. Three individuals were sampled in each group at 3, 7 and 12 d during exposure, and the body wall was frozen for RNA extraction.
2.2
Cloning of the full-length cDNA of CYP4 from M. sanguinea
2.2.1
Degenerate primers
Primer Premier 5.0 software was used for designing primers. Two degenerate primers (F1 and R1) for MsCYP4 were designed using CDS conserved regions from known polychaete P. aibuhitensis gene (GenBank: HM126463) which had been compared with other invertebrates. Gene-specific primers (F2, R2, F3, and R3; Table 1) were subsequently designed on the basis of the obtained cDNA partial sequence.
The individual total RNA was extracted from the pieces of body wall in the first third of M. sanguinea using RNAiso Plus (Takara, Shiga, Japan) according to the manufacturer’s instructions and purified using Recombinant DNase I. Approximately 500 ng/μL RNA was used as template, and the first-strand cDNA synthesis was carried out using TaKaRa RNA PCR Kit (AMV) Ver. 3.0. Subsequently, cDNA was diluted and stored at –20°C.
2.2.3
cDNA cloning
Polymerase chain reaction (PCR) was conducted to obtain the MsCYP4 fragments. F1 and R1 were used in a 25 μL of reaction volume containing 5× PCR buffer (5 μL), primers (10 μmol/L, 0.5 μL each), cDNA (5 μL), TaKaRa Ex Taq HS (0.15 μL), and PCR-grade water (13.85 μL). The PCR program was set at 94°C for 5 min, followed by 30 cycles of 94°C for 30 s, 51°C for 30 s, 72°C for 30 s, and the final extension step at 72°C for 10 min. The PCR products were gel-purified, cloned into the pMDTM-18 T vector (TaKaRa, Shiga, Japan), and transformed into the Escherichia coli competent cell DH5α (TaKaRa, Shiga, Japan). Positive recombinants were identified via blue-white color selection in ampicillin-containing LB plates and sequenced by Sangon Technology Co. Ltd. (Shanghai, China).
2.2.4
Rapid amplification of cDNA 3′ and 5′ ends (3′ and 5′ RACE)
To obtain the full-length MsCYP4 cDNAs, the 5’and 3’RACE PCR technique was used. Four gene-specific primers (F2, R2, F3 and R3; Table 1) were designed on the basis of the fragments obtained from homology cloning.
RNA was reverse-transcribed to first-strand cDNA using 3′-Full RACE Core Set Ver. 2.0. The 3′ end of CYP4 cDNA was amplified using TaKaRa LA Taq® with GC Buffer via nested PCR. In the first round of nested PCR, F2 and UPM-L were used as primers. The reaction volume was 50 μL containing 2 μL of cDNA, 8 μL of 1×cDNA Dilution Buffer II, 2 μL of each 10 mmol/L primer, 25 μL of 2×GC Buffer I, 0.5 μL of TaKaRa LA Taq® (5 U/μL) and 10.5 μL of PCR-grade water. The reaction condition was as follows: 94°C for 3 min, followed by 30 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 2 min, and a final extension at 72°C for 10 min. In the second round of nested PCR, F3 and UPM-S primer were used as primers. The reaction volume was 50 μL containing 1 μL of outer PCR product, 8 μL dNTP (2.5mM each), 2 μL of each 10 mmol/L primer, 25 μL of 2×GC Buffer I, 0.5 μL of TaKaRa LA Taq® (5 U/μL) and 11.5 μL of PCR-grade water. The amplification profile was 94°C for 3 min, followed by 30 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 2 min, and a final extension at 72°C for 10 min.
The 5′ end of CYP4 cDNA was synthesized through nested PCR using 5′-Full RACE Kit (Takara, Shiga, Japan). After Alkaline phosphatase (CIAP) and TAP treatment, RNA was ligated using the 5′-RACE adaptor and reversed to first-strand cDNA. In the first round of nested PCR, R2 and UPM-L were used as primers. The reaction volume was same as 3′-RACE outer PCR except the use of primers and 5′ cDNA instead of 3′ cDNA. The reaction condition was as follows: 94°C for 3 min, followed by 30 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 2 min, and a final extension at 72°C for 10 min. In the second round of nested PCR, R3 and UPM-S were used as primers. The reaction volume was same as 3′-RACE inner PCR except the use of primers and 5′ cDNA instead of 3′ cDNA. The amplification profile was 94°C for 3 min, followed by 30 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 2 min, and a final extension at 72°C for 10 min.
2.2.5
Homology analysis
The resulting sequences were analyzed for similarity with other known sequences using the BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/). The deduced amino acid sequence was analyzed using the Expert Protein Analysis System (http://www.us.expasy.org/tools). Alignment of multiple sequences was also analyzed using the CLUSTAL W (http://www.ebi.ac.uk/clustalW). The functional sites and motifs in amino acid sequences were predicted using Motif Scan (http://www.hits.isbsib.ch/cgi-bin/PESCAN) and Expasy (http://www.au.expasy.org/prosite/). Phylogenetic analysis was conducted via Mega 5.0 software using the neighbor-joining algorithm.
2.3
Quantitative real-time PCR analysis
The total RNA in each treatment was extracted from the body wall using TaKaRa RNAisoTM Plus kit and reverse-transcribed into single-strand cDNA using PrimeScript® RT reagent Kit with gDNA Eraser (Perfect Real Time) (Takara, Shiga, Japan). cDNA was stored at –20°C.
Gene-specific primers for MsCYP4 (F4, R4) were designed for real-time PCR using the Primer Premier 5.0 software. The β-actin housekeeping gene was used as reference gene for internal standardization. The total amplification volume was 20 μL, which included SYBR® Premix Ex TaqTM Ⅱ (10 μL), forward and reverse primer (0.8 μL each), ROX reference dye (0.4 μL), 1:10 diluted cDNA (2 μL), and DEPC-treated water (6 μL). The PCR protocol was 95°C, for 30 s followed by 40 cycles of 95°C for 5 s and 60°C for 30 s. Each amplification product generated a single peak in the dissociation curve analysis, which demonstrated the specificity of the PCR products.
2.4
Statistical analysis
The 2–△△CT method was used to analyze the expression level of mRNA from M. sanguinea. All data were expressed as means±SE. Differences among groups were analysed by one-way ANOVA and Duncan’s multiple range Test using the SPSS 18 at a significance level of p< 0.05.
3.
Results
3.1
Identification of MsCYP4 full-length cDNA
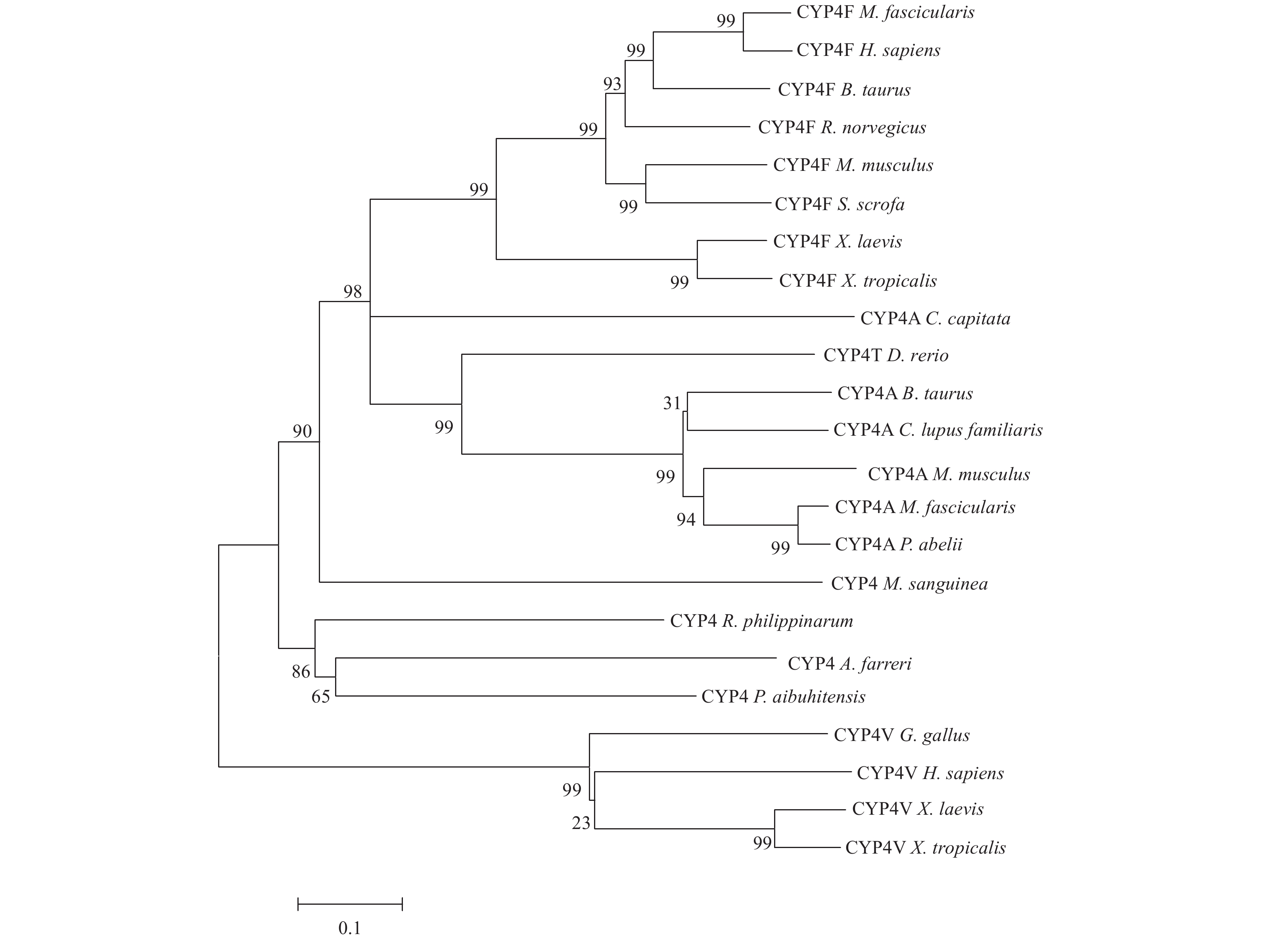
The full-length cDNA of MsCYP4 contains 2 470 nucleotides, including a 3′ untranslated region (UTR, 822 bp), 5′ UTR (109 bp), and an open reading frame of 1 539 nucleotides that is predicted to encode a protein of 512 amino acids with predicted molecular mass of 59.36 kDa and theoretical isoelectric point of 8.12 (Fig. 1). The sequence of M. sanguineaCYP4 cDNA is named MsCYP4 and submitted to NCBI GenBank (GenBank ID: Kf203131). Conserved structural patterns are included in this deduced amino acid sequence. The CYP4-conserved cysteine heme-binding domain, FxxGxRxCxG, is located at the amino acid residues 453–466 aa. The conserved motif ExxR within the K-helix and WxxxR within the C-helix are presented at the amino acid residues 379–382 aa and 141–145 aa, respctively. The CYP4 family characteristic sequence of 13-residues EVDTFMFEGHDTT in the I-helix with the function of proton transfer during monooxygenation. The deduced amino acid sequence shared similarity with known CYP4 enzymes and was highly homology to CYP4F (47%) from Xenopus tropicalis and other CYP family 4 (42%–45%) from invertebrate and vertebrate (Fig. 2). Marphysasanguinea CYP sequence belonged to the CYP family 4 due to the p450 domain (272–1582 aa). Phylogenetic tree was constructed to demonstrate the relationship of this deduced amino acid sequence with other CYP proteins (Fig. 3).
Figure
1.
Full-length cDNA and deduced amino acid sequences of Marphysa sanguinea CYP4. The start and stop codons are shown in red. A heme-binding domain (FxxGxxxCxG) is shadowed, C-helix motif is boxed in black, K-helix motif is boxed in red, characteristic conserved domain of CYP4 EVDTFMFEGHDTT is underlined.
Figure
2.
Multiple alignment of CYP4 protein. Amino acid residues that are conserved in at least 50% sequence are shaded, and similar amino acids are shaded in dark, including Homo sapiens (AAH35350.1), Rattus norvegicus (NP_062569.2), Monodelphis domestica (XP_001367719.1), Xenopus tropicalis (NP_001015810.1), Capitella capitata (AAS87604.1), Saccoglossus kowalevskii (XP_002730972.1), Marphysa sanguinea (HM126463), Chlamys farreri (ACL80141.1), Venerupis philippinarum (ACM16804.2), Perinereis aibuhitensis (ADI52567.1), and Nereis virens (AAR88241.2).
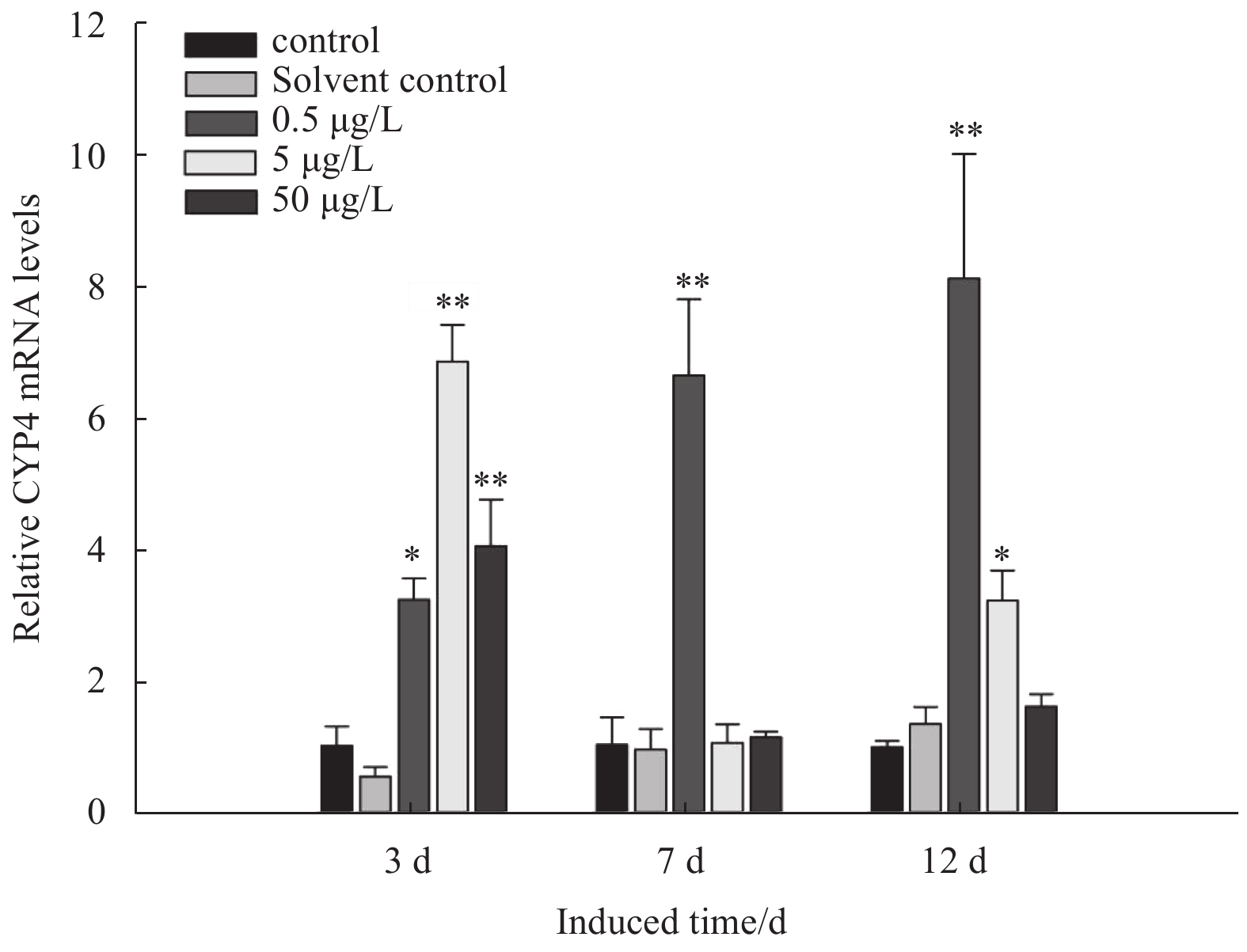
The dose-dependent pattern of MsCYP4 in response to BaP exposure was examined (Fig. 4). Solvent control group showed similar response with blank control group, and no significant difference was observed, which indicated that acetone had no influence on M. sanguinea as a solvent. The transcripts in M. sanguinea exposed to 0.5 μg/L BaP were increased gradually during the exposure period and significantly upregulated to 2.92-, 6.73-, and 7.99-fold, which showed a significant difference with the control (p<0.01). The maximal response was observed at Day 3 in 5 and 50 μg/L, and the mRNA expression levels in 5 and 50 μg/L concentrations were 6.46- and 3.33-fold of those in control which showed significant difference (p<0.01). Dropping abruptly at Day 7, the mRNA expression levels were 1.26- and 1.18-fold of those in control and showed no significant difference (p>0.05). The transcripts of MsCYP4 gene increased slightly by 2.98- and 1.54-fold at Day 12 in 5 and 50 μg/L groups, respectively.
Figure
4.
Relative expression of CYP4 mRNA under benzo[a]pyrene exposure in M. sanguinea. Samples were treated with different concentrations (0.5, 5 and 50 μg/L) for 12 d. * Significant differences among concentrations within sampling times (p<0.05); ** extremely significant differences among concentrations within sampling times (p<0.01).
CYP4 enzymes play an important role in oxidative metabolism of endogenous and exogenous chemicals. These enzymes have diverse functions and structures in both vertebrates and invertebrates, and take on critical roles in phase I detoxification systems (Baldwin et al., 2009). CYP4 can be induced by PAHs and its metabolites in polychaetes (Zheng et al., 2013). Inhabiting in sediment M. sanguinea can accumulate and digested much of PAHs component. Thus, they are widely used as biomarkers in detecting marine pollution caused by PAHs (Han et al., 2014). Here, to understanding the relationship between PAHs and CYP4 activity in M. sanguinea, the MsCYP4 gene was cloned, and the expression pattern was detected.
In the current study, the full-length cDNA of MsCYP4 gene was identified and characterized for the first time. This sequence contains 2 470 bp that encodes 512 amino acids. The deduced amino acid sequence shares a number of common characteristics with other members of the CYP4 family: heme-binding domain (FxxGxxxCxG) serving as the fifth ligand to heme iron (Position 453–466) (Xu et al., 2015), and cysteine residue playing an important part in the structure of binding oxygen; highly conserved domain WxxxR (Positions 141–145) in the C-helix at N-terminal, which can interact with hemoglobin (Liu et al., 2010); 13-residue (EVDTFMFEGHDTT) domain in the I-helix of CYP4 family, which is involved in substrate oxidation reaction and proton transfer (Positions 322–327) (Pan et al., 2011); and conserved domain EXXR (Positions 379-382) in the K-helix, which stabilize the heme core structure (Chen et al., 2014). The multiple sequence alignment analysis showed that the CYP4 amino acid sequence of M. sanguinea had similarity with CYP4s in other species. Numerous alignments corresponding to high 40% similarity confirmed that this M. sanguinea sequence belongs to the CYP4 family (Zheng et al., 2013). This result was in accordance with the findings of Rewitz et al. (2004).
Deposit-feeding invertebrates, such as polychaetes, can be sensitive to the biotransformation of pollutants in sediments. Experiment showed that after the excrement of M. sanguinea had stood on the sediment for 2 h, the PAHs concentrations decrease to half, and this reduction may be caused by microorganisms′chemical changes or enzyme changes in the excrement of M. sanguinea (Onozato et al., 2012). Bach et al. (2005) found 20% of total PAHs to be polar metabolites in Capitella sp. I after 15 d exposure to 30 (μg fluoranthene)/(g sediment), however, Capitella sp. S only 3% of total fluoranthene were present as polar metabolites. The PAHs tolerance was different between two different Capitella species.
Jørgensen et al. (2008) indicated that the biotransformation process of PAHs was similar in both vertebrates and invertebrates which included two stages. In the first stage, CYP enzymes catalyze PAHs into water-soluble metabolites. The second stage is glutathione S-transferase detoxification. Many studies have investigated the induction after exposure to PAHs in marine animals. PAHs have upregulation effects on the mRNA expression of CYP4 genes in polychaete N. virens, C. capitate, and P. nuntia (Nelson, 2011; Ohtsuki et al., 2012; Won et al., 2013; Zheng et al., 2013). The changes indicated that the CYP4 gene expression in polychaete was inducible by PAHs. Miao et al. (2011) showed that exposure to BaP, CfCYP4 was significantly decreased in the gill and digestive gland of scallops. Nelson (2011) proposed that two CYP4 genes in fungi have low expression level under PAHs exposure, and the low expression level may be caused by the low control ability of CYP4 enzymes. In the present study, MsCYP4 mRNA expression level was also induced under BaP exposure. The transcripts of MsCYP4 mRNA increased with exposure time in 0.5 μg/L dose group, and a 7.99-fold induction of MsCYP4 mRNA expression was observed after 12 d. The MsCYP4 mRNA expression firstly showed an increasing trend, then decreased, and finally increased slightly in 5 and 50 μg/L dose group. These different expression trends with other species may be related to the polychaete tolerance of PAHs. The MsCYP4 mRNA expression was increased within the tolerance of the PAHs which explained the rising trend of low concentration groups. The downward trend of MsCYP4 mRNA expression may result from the excessive DNA damage under high concentration of BaP exposure or toxicity effect. Many researchers refer to this phenomenon as induced saturation, that is, when the inducer reached a certain concentration, the gene expression levels would not increase but decrease (Fisher et al., 2003).
Pan et al. (2011) studied the CYP4 mRNA expression in different tissues and showed no notable change in CYP4 mRNA expression in gill of clam R. philippinarum exposed to BaP; on the contrary, the CYP4 mRNA expression was induced significantly by 0.2 μg/L BaP (p<0.05) in the digestive gland of the clam. The different expression levels may be caused by the sampling tissues the experiment used. In the current study, only body wall was used as sample, and further experiment about CYP4 mRNA expression in different tissues is needed. Moreover, the detoxification mechanism of CYP4 in polychaete should be further studied.
In conclusion, we cloned the full length of CYP4 cDNA from M. sanguinea and examined the mRNA expression of MsCYP4 under BaP exposure. The deduced amino acid sequence shared high homology with other proteins in the CYP4 family. Our results proposed a biomarker to detect PAHs pollution in marine environment and indicated that M. sanguinea may be a promising species that can detect coastal environment pollution.
Alam M A, Gomes A, Sarkar S K, et al. 2010. Trace metal bioaccumulation by soft-bottom polychaetes (Annelida) of Sundarban mangrove wetland, India and their potential use as contamination indicator. Bulletin of Environmental Contamination and Toxicology, 85(5): 492–496
Bach L, Palmqvist A, Rasmussen L J, et al. 2005. Differences in PAH tolerance between Capitella species: underlying biochemical mechanisms. Aquatic Toxicology, 74(4): 307–319
Baldwin W S, Marko P B, Nelson D R. 2009. The cytochrome P450 (CYP) gene superfamily in Daphnia pulex. BMC Genomics, 10: 169
Barakat A O, Mostafa A, Wade T L, et al. 2011. Distribution and characteristics of PAHs in sediments from the Mediterranean coastal environment of Egypt. Marine Pollution Bulletin, 62(9): 1969–1978
Chen Wanping, Lee M K, Jefcoate C, et al. 2014. Fungal cytochrome p450 monooxygenases: their distribution, structure, functions, family expansion, and evolutionary origin. Genome Biology and Evolution, 6(7): 1620–1634
Chen Xue, Zhou Yibing, Yang Dazuo, et al. 2012. CYP4 mRNA expression in marine polychaete Perinereis aibuhitensis in response to petroleum hydrocarbon and deltamethrin. Marine Pollution Bulletin, 64(9): 1782–1788
Dean H K. 2008. The use of polychaetes (Annelida) as indicator species of marine pollution: a review. Revista de Biologia Tropical, 56(S4): 11–38
Feng Chenglian, Lei Bingli, Wang Zijian. 2009. Preliminary ecological risk assessment of polycyclic aromatic hydrocarbons in main rivers of China. China Environmental Science (in Chinese), 29(6): 583–588
Fisher T, Crane M, Callaghan A. 2003. Induction of cytochrome P-450 activity in individual Chironomus riparius Meigen larvae exposed to xenobiotics. Ecotoxicology and Environmental Safety, 54(1): 1–6
Guo Hui, Xian Jian’an, Li Bin, et al. 2013. Gene expression of apoptosis-related genes, stress protein and antioxidant enzymes in hemocytes of white shrimp Litopenaeus vannamei under nitrite stress. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 157(4): 366–371
Han J, Won E J, Hwang D S, et al. 2014. Crude oil exposure results in oxidative stress-mediated dysfunctional development and reproduction in the copepod Tigriopus japonicus and modulates expression of cytochrome P450 (CYP) genes. Aquatic Toxicology, 152: 308–317
Hong S, Khim J S, Ryu J, et al. 2012. Two years after the HebeiSpirit oil spill: residual crude-derived hydrocarbons and potential AhR-mediated activities in coastal sediments. Environmental Science & Technology, 46(3): 1406–1414
Jernelöv A. 2010. The threats from oil spills: Now, then, and in the future. Ambio, 39(5–6): 353–366
Jørgensen A, Giessing A M B, Rasmussen L J, et al. 2008. Biotransformation of polycyclic aromatic hydrocarbons in marine polychaetes. Marine Environmental Research, 65(2): 171–186
Jørgensen A, Rasmussen L J, Andersen O. 2005. Characterisation of two novel CYP4 genes from the marine polychaete Nereis virens and their involvement in pyrene hydroxylase activity. Biochemical and Biophysical Research Communications, 336(3): 890–897
Kirischian N L, Wilson J Y. 2012. Phylogenetic and functional analyses of the cytochrome P450 family 4. Molecular Phylogenetics and Evolution, 62(1): 458–471
Kraugerud M, Doughty R W, Lyche J L, et al. 2012. Natural mixtures of persistent organic pollutants (POPs) suppress ovarian follicle development, liver vitellogenin immunostaining and hepatocyte proliferation in female zebrafish (Danio rerio). Aquatic Toxicology, 116–117: 16–23
Lewis C, Watson G J. 2012. Expanding the ecotoxicological toolbox: the inclusion of polychaete reproductive endpoints. Marine Environmental Research, 75: 10–22
Liu Fang, Jiang Hongling, Ye Songqing, et al. 2010. The Arabidopsis P450 protein CYP82C2 modulates jasmonate-induced root growth inhibition, defense gene expression and indole glucosinolate biosynthesis. Cell Research, 20(5): 539–552
Lyche J L, Grześ I M, Karlsson C, et al. 2013. Parental exposure to natural mixtures of POPs reduced embryo production and altered gene transcription in zebrafish embryos. Aquatic Toxicology, 126: 424–434
Lyche J L, Nourizadeh-Lillabadi R, Karlsson C, et al. 2011. Natural mixtures of POPs affected body weight gain and induced transcription of genes involved in weight regulation and insulin signaling. Aquatic Toxicology, 102(3–4): 197–204
Martins M, Costa P M, Ferreira A M, et al. 2013. Comparative DNA damage and oxidative effects of carcinogenic and non-carcinogenic sediment-bound PAHs in the gills of a bivalve. Aquatic Toxicology, 142–143: 85–95
Miao Jingjing, Pan Luqing, Liu Na, et al. 2011. Molecular cloning of CYP4 and GSTpi homologues in the scallop Chlamys farreri and its expression in response to benzo[a]pyrene exposure. Marine Genomics, 4(2): 99–108
Musale A S, Desai D V. 2011. Distribution and abundance of macrobenthic polychaetes along the South Indian coast. Environmental Monitoring and Assessment, 178(1–4): 423–436
Nelson D R. 2011. Progress in tracing the evolutionary paths of cytochrome P450. Biochimica et Biophysica Acta (BBA)—Proteins and Proteomics, 1814(1): 14–18
Ohtsuki S, Schaefer O, Kawakami H, et al. 2012. Simultaneous absolute protein quantification of transporters, cytochromes P450, and UDP-glucuronosyltransferases as a novel approach for the characterization of individual human liver: comparison with mRNA levels and activities. Drug Metabolism and Disposition, 40(1): 83–92
Onozato M, Sugawara T, Nishigaki A, et al. 2012. Study on the Degradation of Polycyclic Aromatic Hydrocarbons (PAHs) in the Excrement of Marphysa sanguinea. Polycyclic Aromatic Compounds, 32(2): 238–247
Pan Luqing, Liu Na, Xu Chaoqun, et al. 2011. Identification of a novel P450 gene belonging to the CYP4 family in the clam Ruditapes philippinarum, and analysis of basal- and benzo(a)pyrene-induced mRNA expression levels in selected tissues. Environmental Toxicology and Pharmacology, 32(3): 390–398
Rewitz K F, Kjellerup C, Jørgensen A, et al. 2004. Identification of two Nereis virens (Annelida: Polychaeta) cytochromes P450 and induction by xenobiotics. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 138(1): 89–96
Scornaienchi M L, Thornton C, Willett K L, et al. 2010. Functional differences in the cytochrome P450 1 family enzymes from Zebrafish (Danio rerio) using heterologously expressed proteins. Archives of Biochemistry and Biophysics, 502(1): 17–22
Sim M S, Jo I J, Song H G. 2010. Acute health problems related to the operation mounted to clean the Hebei Spirit oil spill in Taean, Korea. Marine Pollution Bulletin, 60(1): 51–57
Song Yingying, Yuan Xiutang, Zhang Shengli, et al. 2011. Single and Joint Toxic Effects of Benzo(a)pyrene and Cadmium on Development of three-setiger Juvenile of Polychaete Pernereis aibuhitensis Grube. Marine Environmental Science (in Chinese), 30(3): 333–336
Tobiszewski M, Namieśnik J. 2012. PAH diagnostic ratios for the identification of pollution emission sources. Environmental Pollution, 162: 110–119
Tsai W T, Mi H H, Chang Yuanming, et al. 2007. Polycyclic aromatic hydrocarbons (PAHs) in bio-crudes from induction-heating pyrolysis of biomass wastes. Bioresource Technology, 98(5): 1133–1137
Uno S, Sakurai K, Nebert D W, et al. 2014. Protective role of cytochrome P450 1A1 (CYP1A1) against benzo[a]pyrene-induced toxicity in mouse aorta. Toxicology, 316: 34–42
Won E J, Rhee J S, Shin K H, et al. 2013. Complete mitochondrial genome of the marine polychaete, Perinereis nuntia (Polychaeta, Nereididae). Mitochondrial DNA, 24(4): 342–343
Xu Jun, Wang Xinyu, Guo Wangzhen. 2015. The cytochrome P450 superfamily: Key players in plant development and defense. Journal of Integrative Agriculture, 14(9): 1673–1686
Zanette J, Goldstone J V, Bainy A C D, et al. 2010. Identification of CYP genes in Mytilus (mussel) and Crassostrea (oyster) species: first approach to the full complement of cytochrome P450 genes in bivalves. Marine Environmental Research, 69(Suppl 1): S1–S3
Zhao Huan, Wang Yixiao, Yang Dazuo, et al. 2016. An analysis of genetic diversity in Marphysa sanguinea from different geographic populations using ISSR polymorphisms. Biochemical Systematics and Ecology, 64: 65–69
Zheng Shenli, Chen Bin, Qiu Xiaoyan, et al. 2013. Three novel cytochrome P450 genes identified in the marine polychaete Perinereis nuntia and their transcriptional response to xenobiotics. Aquatic Toxicology, 134–135: 11–22
Zhou Chi, Li Chunhou, Zhang Weimin, et al. 2010a. CYP4 gene cloning and expression level analysis of Perna viridis. Journal of Tropical Oceanography (in Chinese), 29(4): 82–88
Zhou Xiaojie, Sheng Changfa, Li Mei, et al. 2010b. Expression responses of nine cytochrome P450 genes to xenobiotics in the cotton bollworm Helicoverpa armigera. Pesticide Biochemistry and Physiology, 97(3): 209–213
O.A. Karlsen, J.D. Rasinger, M. Brattås, et al. Quantitative analysis of the polar cod (Boreogadus saida) hepatic proteome highlights interconnected responses in cellular adaptation and defence mechanisms after dietary benzo[a]pyrene exposure. Science of The Total Environment, 2025, 963: 178510. doi:10.1016/j.scitotenv.2025.178510
2.
Amarachi P. Onyena, Cathrine S. Manohar, Joseph A. Nkwoji, et al. Characterization of the molecular differential responses in marine benthic macroinvertebrates exposed to polycyclic aromatic hydrocarbons. Aquatic Ecology, 2024, 58(2): 263. doi:10.1007/s10452-023-10064-4
3.
Jiabo He, Zhi Chen, Chen Jing, et al. Behavioral and biochemical responses of the marine polychaete Perinereis aibuhitensis to 2-ethylhexyl-4-methoxycinnamate (EHMC) exposure. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2024, 279: 109868. doi:10.1016/j.cbpc.2024.109868
4.
Alan J. Mearns, Ann Michelle Morrison, Courtney Arthur, et al. Effects of pollution on marine organisms. Water Environment Research, 2020, 92(10): 1510. doi:10.1002/wer.1400
5.
Zhenzhen Zhou, Bin Wang, Shanmei Zeng, et al. Glutathione S-transferase (GST) genes from marine copepods Acartia tonsa: cDNA cloning and mRNA expression in response to 1, 2-dimethylnaphthalene. Aquatic Toxicology, 2020, 224: 105480. doi:10.1016/j.aquatox.2020.105480
6.
Maria Claudia R. Sola, Aldenor G. Santos, Sabrina T. Martinez, et al. Occurrence of 3-nitrobenzanthrone and other powerful mutagenic polycyclic aromatic compounds in living organisms: polychaetes. Scientific Reports, 2020, 10(1) doi:10.1038/s41598-020-60369-9
7.
Donglei Wu, Zhiquan Liu, Mingqi Cai, et al. Molecular characterisation of cytochrome P450 enzymes in waterflea (Daphnia pulex) and their expression regulation by polystyrene nanoplastics. Aquatic Toxicology, 2019, 217: 105350. doi:10.1016/j.aquatox.2019.105350
Figure 1. Full-length cDNA and deduced amino acid sequences of Marphysa sanguinea CYP4. The start and stop codons are shown in red. A heme-binding domain (FxxGxxxCxG) is shadowed, C-helix motif is boxed in black, K-helix motif is boxed in red, characteristic conserved domain of CYP4 EVDTFMFEGHDTT is underlined.
Figure 2. Multiple alignment of CYP4 protein. Amino acid residues that are conserved in at least 50% sequence are shaded, and similar amino acids are shaded in dark, including Homo sapiens (AAH35350.1), Rattus norvegicus (NP_062569.2), Monodelphis domestica (XP_001367719.1), Xenopus tropicalis (NP_001015810.1), Capitella capitata (AAS87604.1), Saccoglossus kowalevskii (XP_002730972.1), Marphysa sanguinea (HM126463), Chlamys farreri (ACL80141.1), Venerupis philippinarum (ACM16804.2), Perinereis aibuhitensis (ADI52567.1), and Nereis virens (AAR88241.2).
Figure 3. Phylogenetic neighbor-joining tree of the 512 amino acid sequences of CYP4 from M. sanguienea with other species. The other CYP4 were as follow: Macaca fascicularis (NP_001270193.1), Homo sapiens (AAH35350.1), Bos taurus (NP_001030214.1), Rattus norvegicus (NP_062569.2), Mus musculus (NP_570952.1), Sus scrofa (NP_001231565.1), Xenopus laevis (NP_001091388.1), Xenopus tropicalis (NP_001015810.1), Capitella capitata (AAS87604.1), Bos taurus (NP_001071376.1), Canis lupus familiaris (NP_001041499.1), Mus musculus (NP_758510.2), Macaca fascicularis (NP_001270811.1), Pongo abelii (NP_001126777.1), Danio rerio (NP_954686.1), Gallus gallus (NP_001001879.1), Xenopus laevis (NP_001086053.1), Xenopus tropicalis (NP_001072667.1), Homo sapiens (EAX04624.1), Azumapecten farreri (ACL80141.1), Ruditapes philippinarum (ACM16804.2), and Perinereis aibuhitensis (ADI52567.1).
Figure 4. Relative expression of CYP4 mRNA under benzo[a]pyrene exposure in M. sanguinea. Samples were treated with different concentrations (0.5, 5 and 50 μg/L) for 12 d. * Significant differences among concentrations within sampling times (p<0.05); ** extremely significant differences among concentrations within sampling times (p<0.01).



 DownLoad:
DownLoad: