Figure
1.
Polyconchoecia chenii sp. nov. (♀). a. Left valve, external view; b. carapace, internal view; and c. postero-ventral corner of right valve, internal view.
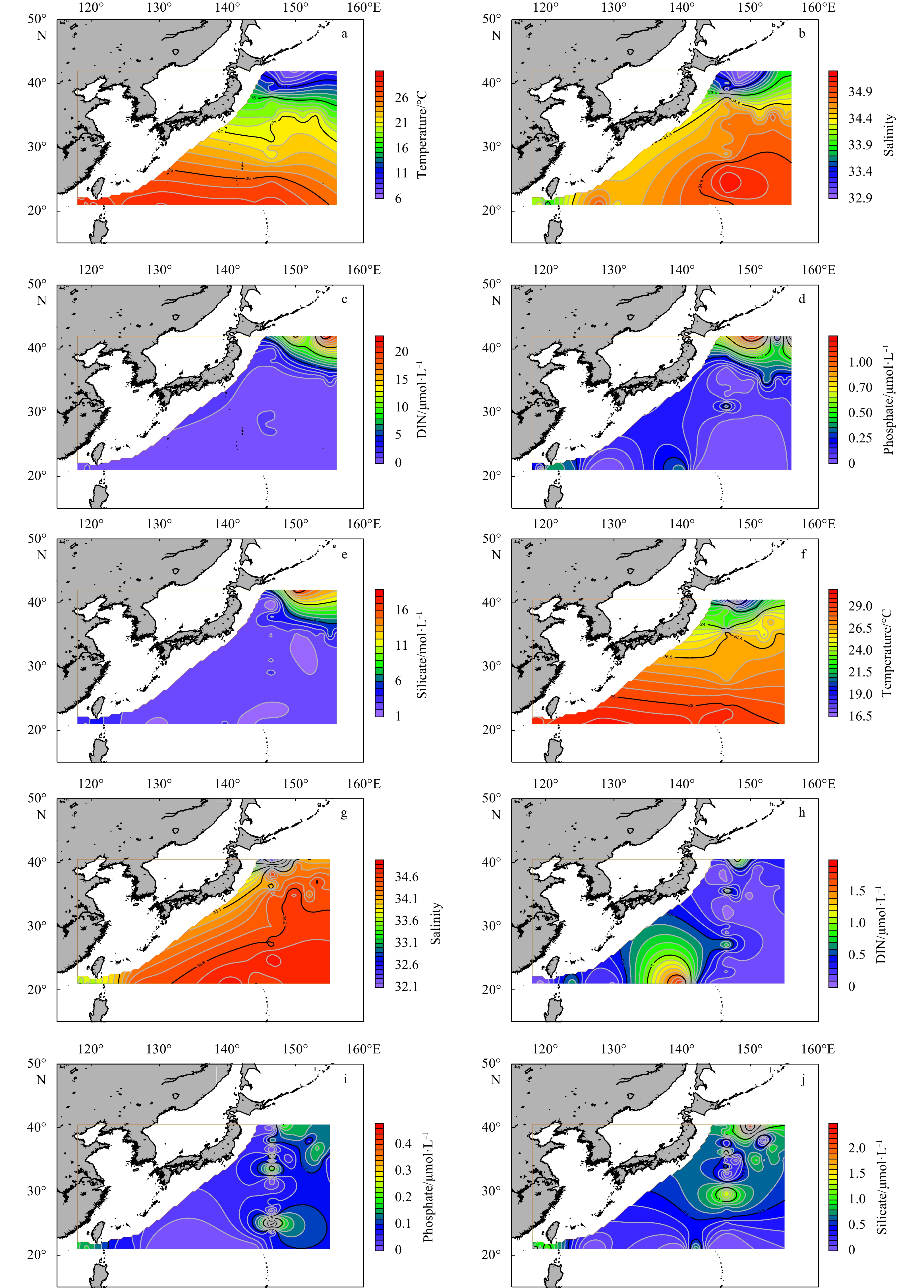
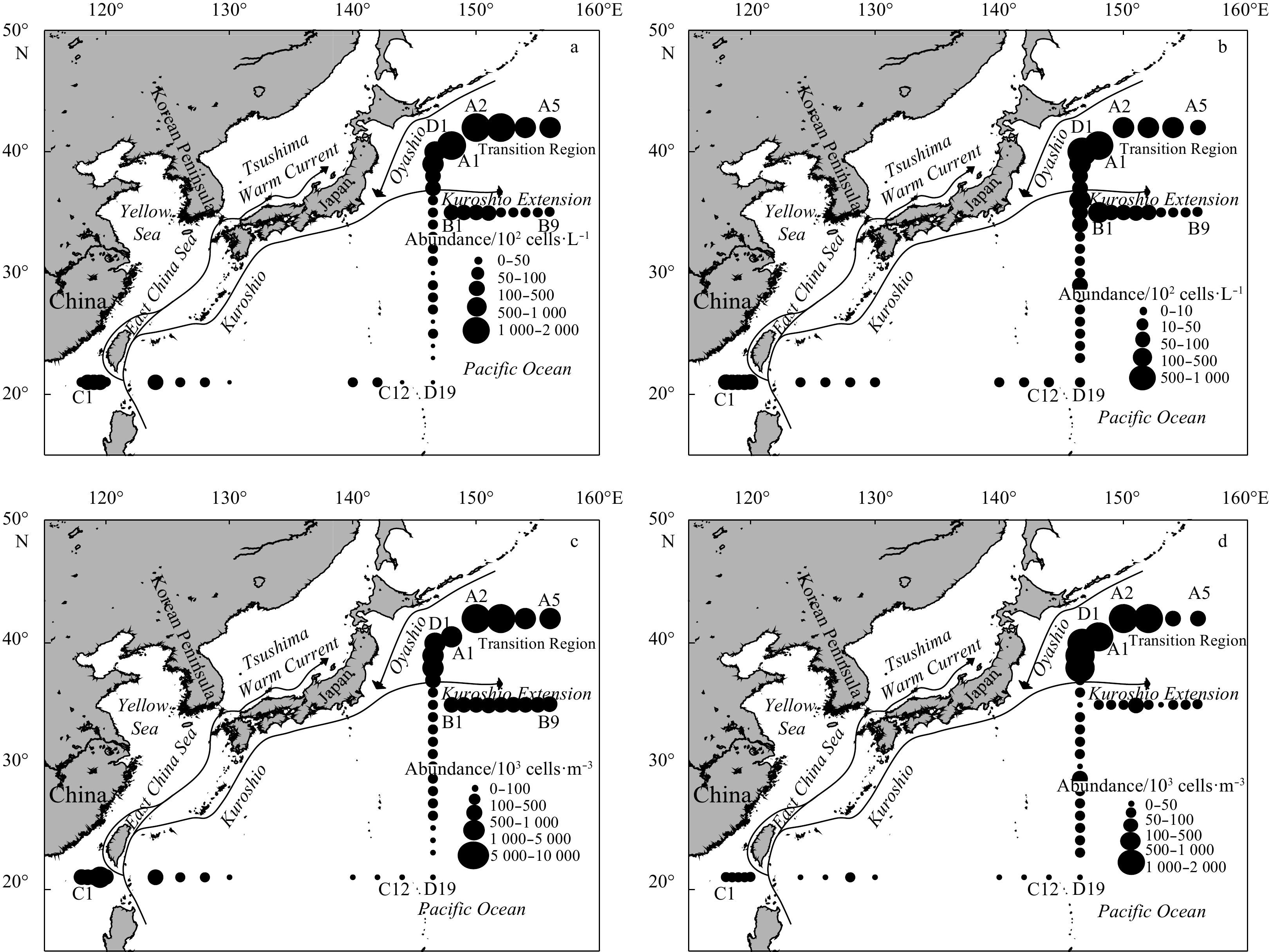
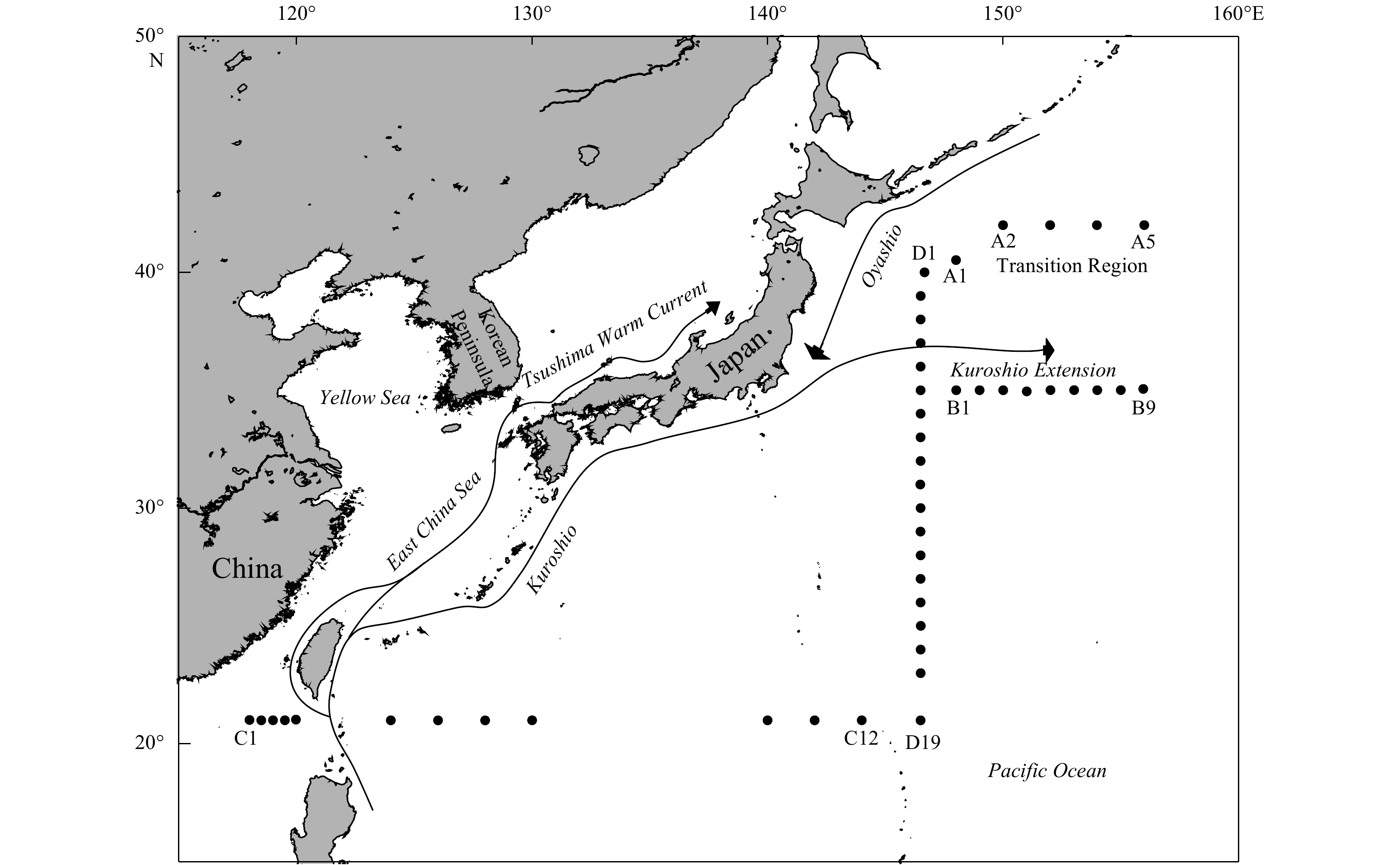
| Citation: | Gengming Lin, Yanghang Chen, Jiang Huang, Yanguo Wang, Youyin Ye, Qingliang Yang. Regional disparities of phytoplankton in relation to different water masses in the Northwest Pacific Ocean during the spring and summer of 2017[J]. Acta Oceanologica Sinica, 2020, 39(6): 107-118. doi: 10.1007/s13131-019-1511-6

|
Class Ostracoda Latreille, 1802, emend. Martin and Davis, 2001 are a big group of small bivalved crustaceans. They inhabit almost all aquatic environments with high taxonomic diversity (Karanovic, 2010). Ostracods are one of the momentous groups of marine organisms, and are functionally important in the process of bio-geo-chemical cycles in marine ecosystem, especially in the tropics and subtropical regions (Angel et al., 2007; George and Nair, 1980). They also play a significant participant of marine deposition, because their calcified valves are preserved as fossils (Di Celma et al., 2016). The studies of ostracods have been well improved in last two decades, and many new taxa have been erected (Harrison-Nelson and Kornicker, 2000; Chavtur, 2003; Lum et al., 2008; Karanovic, 2010; Chavtur and Angel, 2011; Pinto and Jocqué, 2013; Xiang et al., 2017a, b, 2018; Du et al., 2018). These faunae might no-monophyletic, phylogeny remains indistinct, and classification is based on typical characteristics (Fortey and Thomas, 1998; Yamaguchi and Endo, 2003).
Halocyprid ostracods constitute appreciable part of marine zooplankton. They are floating through virtually everywhere in marine environment including surface, hydrothermal vent, cold seep and abyss (van Harten, 1992; Oakley et al., 2013; Tanaka and Yasuhara, 2016; Yamaguchi et al., 2016). In 1853, the largest family Halocyprididae Dana, 1853 (Chavtur, 2003; Martin and Davis, 2001; Brandão et al., 2019) within order Halocyprida Dana, 1853 was erected. And then, the subfamily Conchoeciinae Müller, 1912 was erected under this family. In 2011, Chavtur and Angel (2011) designated the tribe Conchoeciini Chavtur and Angel, 2011 on the basis of the locational shifts of glands. Not long ago, Du et al. (2018) erected genus Polyconchoecia Xiang, Chen and Du, 2018 with the type species P. commixtus Xiang, Chen and Du, 2018 based on the definite characteristics of locations of major glands on carapace. Soon after that, genus Conchoecia Dana, 1849 was subdivided into five genera: Conchoecia; Macrochoecilla Chavtur, 2018; Lophuroecia Chavtur, 2018; Parvidentoecia Chavtur, 2018; and Hyalocoecia Chavtur, 2018; and genus Parthenoecia Chavtur, 2018 was erected, mainly based on characteristics of lateral gland, armature of seta-e on the first antenna in the male, and copulatory appendage (Chavtur and Bashmanov, 2018). Thus far, the tribe Conchoeciini has contained 27 genera (Brandão et al., 2019).
In this study, a new species of genus Polyconchoecia from the South China Sea was described.
Collections were obtained from two cruises of the South China Sea in 2014–2015. All zooplankton specimens were collected using a Multinet sampling system (Type Midi, Mesh-size aperture 200 µm, HydroBios Inc., Kiel, Germany) by vertical and stratified hauls from bottom to surface. Collections were preserved by immersion in 5% buffered formaldehyde.
Specimens were dissected under a Carl Zeiss Discovery V20 zoom-stereomicroscope. Dissected appendages were mounted in permanent slides with CMC-9AF medium (Masters Company Inc., Illinois, USA). Observations were done by a Carl Zeiss Axio Imager Z2 differential interference contrast microscope system with AxioVision Image-Pro software (Carl Zeiss Inc., Oberkochen, Germany). All drawings were made from micro-images of dissected appendages, followed the methodology in Chavtur and Angel (2011), and further processed with Adobe Photoshop CS6 (Adobe Inc., San Jose, CA, USA).
The type specimens/appendages were deposited in the Marine Biological Sample Museum of the Chinese Offshore Investigation and Assessment, Third Institute of Oceanography, MNR (Xiamen, China), under the collection numbers TIO-OHH-PP-201 to TIO-OHH-PP-204.
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:C8D3334A-AA1C-4355-AB53-140B299051B0. The electronic edition of this work was published in a journal with an ISSN, and has been archived and is available from the following digital repositories: SpringerLink, PubMed Central, LOCKSS.
Oder Halocyprida Dana, 1853
Family Halocyprididae Dana, 1853
Subfamily Conchoeciinae Müller, 1912
Tribe Conchoeciini Chavtur and Angel, 2011
Genus Polyconchoecia Xiang, Chen and Du, 2018
Species Polyconchoecia chenii Xiang, Wang and Chen sp. nov.







LSID: urn:lsid:zoobank.org:act:9B60314F-EE85-46C7-8213-737601ED74D8
Etymology. Latinized name of Ruixiang Chen, our teacher, a scientist of the planktonic research group, Third Institute of Oceanography, MNR, in recognition of his important contributions of marine ostracods of China.
Holotype. No. TIO-OHH-PP-201, adult female, length 2.57 mm, height 1.47 mm from Sta. CS-068 (14°31′N, 114°54′E) in the top of the South China Sea, 200–500 m water layer, 12 January 2015. Specimen was dissected on slide and deposited in the Marine Biological Sample Museum, in the Third Institute of Oceanography, MNR, China (Xiamen, China).
Paratypes. No. TIO-OHH-PP-202, adult female, length 2.33 mm, height 1.35 mm, No. TIO-OHH-PP-203, adult female, length 2.45 mm, height 1.41 mm, from the same locality of the holotype, and dissected on slide. TIO-OHH-PP-204, adult female, length 2.35 mm, height 1.33 mm, collected from Sta. CS-012 (7°06.46′N, 113°53.6′E) in the South China Sea, 200–500 m water layer on 2 January 2014, deposited in 5% buffered formaldehyde. Paratypes were deposited with the holotype.
Distribution. The mesopelagic water in the South China Sea.
Diagnosis. Carapace without ornamentation or setae, height about 56.5%–57.9% of length, sub-rectangle in lateral view with rounded corners; rostrums wide, developed, equilong, anteriorly and curved to downward; shoulder vaults unconspicuous, higher in anterior part, ventral margin with slightly concave, left asymmetric gland opening on postero-dorsal corner, a lateral gland opening on right postero-ventral corner, dense edge glands placed along all ventral margin; left postero-ventral margin without gland. Frontal organ segmented. In first antenna, a- and c-setae analogic with long end joint, b- and d- setae analogic without end joint. E-seta of second antenna present. Maxilla with five disto-anterior setae, one disto-medial seta, two medial setae, and three disto-posterior setae on endopod 2. Proximal-ventral group of setae of fifth limb with seven setae, endopod 1 with two ventral setae. Vesting of exopod of sixth limb strong and spinose, basale with seven ventral plumose setae, endopod 1 bare, endopod 2 without ventral seta. Furca with unpaired seta.
Carapace (Figs 1a-c and 7a, b). Carapace smooth without setae and ornamentation (pits or grooves), sub-rectangle in lateral view; shoulder vaults unconspicuous; rostrum subequal anteriorly with acutangular tip, curved downward, wide and developed; antero-ventral margin, postero-dorsal and postero-ventral corner rounded; anterior part slightly higher than posterior part; dorsal margin approximately flat; ventral margin with slightly concavity. Carapace with three groups of glands: one asymmetric gland opening on postero-dorsal corner of left valve, one lateral gland opening on right postero-ventral corner; dense edge glands placed along anterior to posterior ventral margin; postero-ventral corner of left valve without gland. Length 2.33–2.57 mm, height 1.33–1.47 mm, height about 56.5%–57.9% of length.
Frontal organ (Figs 2a, c and 7c, d). Stem and capitulum separated, straight and clavate with blunt tip, small disto-dorsal and ventral spines. Capitate base separated from first antenna.
First antenna (Figs 2a, b, d and 7c). First antenna uniramous. Basale and endopod 1 bare. Endopod 2 with one small dorsal spinose seta. Endopod 3–5 very short and small. Endopod 4 with two ventral sensory setae (a- and b-setae). Endopod 5 blunt conical with three disto-ventral sensory setae (c-, d-, e-setae). A- and c-setae analogic long columnar, thin walled and bare with long end joint; b- and d- setae equilong and bare without end joint, slightly longer than a- and c-setae; e-seta extremely long (approximately one and half length of a-seta) and spinose ventral spines with numerous small ventral spines on distal half.
Second antenna (Figs 2e, f, 3a, b and 7e, f). Limb biramous with large protopodite with powerful muscles. Endopod without c- and d-setae. Endopod 1 large, folded forward, with bend a- and b-setae, a-seta bare and short, b-seta long and spinose, about twice length of a-seta; processus mamillaris normal. Endopod 2 and 3 integrated into a small peg shaped bulge on disto-dorsal margin of endopod 1, with bare 5 setae; g-seta longest and ringed; f-seta second longest; h-, i-, j- setae equilong. Exopod 1 more than seventeen times length of 2, with one bend acerose spine on disto-dorsal margin instead of plumose seta; exopod 2–7 very short with analogous long plumose swimming seta on disto-ventral margin respectively; exopod 8 and 9 fused with one long plumose seta, one shorter plumose seta and one very small seta on tip.
Mandible (Figs 3c-e, 4a-c and 7g-j). Basale large. Exopod tiny peg shaped, with one dorsal plumose seta with bare distal part. Endopod 1 more than two times length of 2, with one disto-ventral seta. Endopod 2 short, with two setae on disto-ventral margin: outer one bare and short, inner one long with small spines on distal half length; three setae on disto-dorsal edge: mid one short and bare, others long with small ventral spines. Endopod 3 very short, with six setae on tip: seta 1 claw-shaped with distal ventral spines; seta 2 small with ventral spines; seta 3 claw-shaped and biggest with disto-ventral spines; setae 4 and 6 subequal in length; seta 5 about twice length of seta 4. Toothed edge of basale big and triangular with two bare long setae on medio-ventral side and two bare short setae on medio-distal side. Toothed edge of basale with eight distal teeth in one list. Coxal endite constituting by three parts: distal teeth list with one big and bend distal triangular papillae, one big papilla constituted four small teeth, five strip papillae, and one flat papilla; proximal teeth list with small teeth; medial part with some long and big papillae and numerous long cilia.
Maxilla (Figs 4d-f and 8a, b). Exopod with two spinose setae on tip. Endopod 1 big rectangular, with two long and three short bare setae on antero-distal edge, one spinose seta on disto-posterior edge, two long spinose setae on medial side, one long and two short spinose setae on posterior side. Terminal segment with five claws: two bilateral stout spinose claws, two inner puny bare claws and one mid spinose claw; spinose claws with disto-ventral spines. Maxilla with three hirsute endites: endite I with five plumose setae; endite II with one bare and three plumose setae; endite III with ten bare setae.
Fifth limb (Figs 5a-c, 6c and 8c). Limb biramous. Basale large and wide with eleven setae: dorsal seta (vestige of exopod) long and bare; proximal-ventral group of setae with five plumose setae and a pair of symmetric ventral spinose setae; disto-ventral group of setae with one central seta and a pair of symmetric ventral spinose setae. Endopod 1 long with two ventral setae and one dorsal seta; endopod 2 short with three long bare curved claws on tip, mid claw longest, mid and ventral claws with numerous small ventral spines. Coxale with three endites: endite I big rectangular with one short bare and one long plumose proximal setae, one small bare one small plumose and one long plumose distal setae; endite II very short with one long plumose seta; ventral group of setae of endite III with two strong bare and blunt setae, and four sharp plumose setae. Epipod with one small bare inner seta and about sixteen lathy lithe plumose setae.
Sixth limb (Figs 5d, 6d and 8d). Basale broad with one long plumose seta (vesting of exopod), two long proximo-ventral, two short ventral and three short disto-ventral plumose setae; basale without dorso-lateral seta. Exopod 1 short with one ventral seta. Exopod 2 long and thin, with one dorsal and one ventral seta. Terminal segment blunt and very short with three very long acerose claws on tip; mid claw longest with curved distal part; mid and ventral claws with numerous small disto-ventral spines. Epipod with one small bare inner seta and about seventeen lathy lithe plumose setae.
Seventh limb (Figs 6a, e and 8e). Segment 1 slender and bare. Segment 2 very short conical with one bare long seta and one bare lathy lithe seta (four times length of another) on tip. Epipod with one small bare inner seta and about fourteen lathy lithe plumose setae.
Furca (Figs 6b and 8f). Each furcal lamella with one large dorsal seta and seven claws, from long to short in turn arrangement. Seta and claws with numerous small disto-ventral spines. Furca with ventral unpaired seta.
These specimens are considered to be one species of the subfamily Conchoeciinae Müller, 1912 of family Halocyprididae Dana, 1853, according to the Chen and Chavtur’s diagnosis (Chen and Lin, 1995; Chavtur and Angel, 2011), and then we can put them into the tribe Conchoeciini, Chavtur and Angel, 2011 easily. They are very close to Polyconchoecia commixtus, the type species of genus Polyconchoecia, which genus has been reported recently and with only one species yet. They have shared characteristics as followed: (1) similar size of full adult; (2) the shapes of carapace are very close; (3) left valve has a left asymmetric gland near postero-dorsal corner; (4) right valve has a lateral gland on right postero-ventral corner; (5) dense edge glands are placed along anterior to posterior ventral margin of carapace in line; (6) exopod 1 of second antenna has one bend acerose spine on disto-dorsal margin instead of plumose seta; (7) the structures of main limbs are similar.
Although these specimens are very close to P. commixtus, P. chenii sp. nov. is discriminated by the morphological comparisons shown in Table 1. They have obvious and individual distinctions to separate from P. commixtus: (1) left asymmetric gland of carapace of the new species is moved posteriorly, the carapace have no right asymmetric gland or compound gland (Figs 1b, c and 7a, b); (2) the frontal organ of this species is separated to stem and capitulum, and has more ventral spines (Figs 2a and 7c, d); (3) in P. chenii sp. nov., the seta of endopod 2 of the first antenna is very small; (4) in P. commixtus, a- to d-setae of the sensory setae of the first antenna are analogic with long end joint, e-seta has short end joint; in these specimens, a- and c-setae are analogic with long end joint, b- and d-setae are analogic without end joint, e-seta has no end joint; (5) in P. commixtus, the endopod of the second antenna has not c-, d- and e-setae, and one small oval hump with central concave on mid-ventral margin, instead of processus mamillaris; in these specimens, e-seta is present and the processus mamillaris is normal (Figs 2f and 7e); (6) in P. commixtus, the exopod 2-4 of the second antenna have no swimming setae (Figs 2e, 3a and 7f); (7) they have unsimilar tooth edge of the coxal endite (Figs 3b, c, 7j and Table 1); (8) they have different setal counts of the mandible, maxilla, fifth limb, and sixth limb (detailed numbers are given in Table 1); (9) P. chenii sp. nov. has unpaired seta on the furcal lamella (Figs 6b and 8f).
| Characteristics | P. commixtus | P. chenii sp. nov. | |
| Carapace | left asymmetric gland | near postero-dorsal margin, moved anteriorly | on postero-dorsal corner |
| right asymmetric gland | on right postero-ventral corner | none | |
| right lateral gland | constituting a compound gland with right asymmetric gland | only on right postero-ventral corner | |
| Frontal organ | stem and capitulum | fused | separated |
| spines | small disto-ventral and mid-ventral spines | small disto-dorsal and ventral spines | |
| First antenna | endopod 2 | with one long dorsal plumose seta | with one small dorsal plumose seta |
| sensory setae | a- to d-setae analogic, long columnar, thin walled and bare, with long end joint; e-seta bare, with short end joint | a- and c-setae analogic, long columnar, thin walled and bare with long end joint; b- and d- setae analogic, equilong and bare without end joint, slightly longer than a- and c-setae; e- seta spinose, without short end joint | |
| Second antenna | endopod | c-, d-, e-setae absent; b-seta bare; one small oval hump with central concave on mid-ventral margin, instead of processus mamillaris | e-setae present; b-seta spinose; processus mamillaris normal. |
| exopod | exopod 1 and 2 fused, exopod 2-4 bare, exopod 8 and 9 separated; terminal plumose seta with single tip | exopod 1 and 2 separated, exopod 2-4 with plumose setae, exopod 8 and 9 fused | |
| Mandible | coxale teeth side | with eight distal teeth | with six distal teeth |
| tooth endites | with four tooth plates | with three tooth plates | |
| endopod | endopod 1 with one ventral seta, and one dorsal seta; endopod 2 with one disto-ventral seta; terminal segment with seven spinose setae | endopod 1 with one ventral seta, without dorsal seta; endopod 2 with two disto-ventral setae; terminal segment with six setae, ventral three bare | |
| Maxilla | endopod | endopod 1 with six disto-anterior setae, one disto-medial seta, none medial seta, and two disto-posterior setae; mid seta of terminal segment bare | endopod 1 with five disto-anterior setae, one disto-medial seta, two medial setae, and three disto-posterior setae; mid seta of terminal segment spinose |
| endites | endite I with eleven plumose setae, endite II with ten plumose setae, endite III with eight plumose setae | endite I with five plumose setae; endite II with one bare and three plumose setae; endite III with ten bare setae | |
| Fifth limb | basale | proximal-ventral group of setae with four setae | proximal-ventral group of setae with seven setae |
| endopod | endopod 1 with one ventral seta | endopod 1 with two ventral setae | |
| endites | endite I with one seta; endite II with two setae; endite III with one proximal seta, three sharp and two blunt distal setae | endite I with five setae; endite II with one seta; endite III with none proximal seta, four sharp and two blunt distal setae | |
| Sixth limb | basale | disto-dorsal seta (vesting of exopod) small and bare; three small ventral setae. | disto-dorsal seta (vesting of exopod) strong and spinose; seven ventral plumose setae |
| endopod | endopod 1 with one ventral seta; endopod 2 with one ventral seta | endopod 1 bare; endopod 2 without ventral seta | |
| Furca | unpaired seta | no | yes |
 DownLoad:
CSV
DownLoad:
CSV
The ostracod faunae of the South China Sea have been known mainly from plankton surveys in surface water or euphotic zone (0–200 m). The local ostracod diversity of deep environments may equal or exceed that of their epi-pelagic relatives (Gianni, 2004; Danovaro et al., 2008). In these years, there are more and more deep-water species have been discovered and reported (Yin et al., 2014; Tanaka and Yasuhara, 2016; Du et al., 2018; Xiang et al., 2018). This work is the second discovery of the genus Polyconchoecia from the world.
We thank Patrick Page-McCaw (Vanderbilt University, USA), Mingyu Li (Xiamen University, China), Guangcheng Chen for critical reading of the manuscript. We are thankful for our zooplankton research group of Third Institute of Oceanography, MNR, for their valuable suggestions for the manuscript preparation.
| [1] |
Allen G R. 2007. Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquatic Conservation Marine and Freshwater Ecosystems, 18(5): 541–556
|
| [2] |
Briggs J C. 2005. The marine East Indies: diversity and speciation. Journal of Biogeography, 32(9): 1517–1522. doi: 10.1111/jbi.2005.32.issue-9
|
| [3] |
Chen C T A, Wang S L. 1999. Carbon, alkalinity and nutrient budgets on the East China Sea continental shelf. Journal of Geophysical Research, 104: 20675–20686. doi: 10.1029/1999JC900055
|
| [4] |
Edmond J, Spivack A, Grant B, et al. 1985. Chemical dynamics of the Changjiang estuary. Continental Shelf Research, 4: 17–36. doi: 10.1016/0278-4343(85)90019-6
|
| [5] |
Egge J K. 1998. Are diatoms poor competitors at low phosphate concentrations?. Journal of Marine Systems, 16: 191–198. doi: 10.1016/S0924-7963(97)00113-9
|
| [6] |
Falkowski P G, Woodhead A D. 1992. Primary Productivity and Biogeochemical Cycles in the Sea, Vol. 43. New York, NY: Plenum Press
|
| [7] |
Fisher T R, Peele E R, Ammerman J W, et al. 1992. Nutrient limitation of phytoplankton in Chesapeake Bay. Marine Ecology Progress Series, 82: 51–63. doi: 10.3354/meps082051
|
| [8] |
Gaston K J. 2000. Global patterns in biodiversity. Nature, 405(6783): 220–227. doi: 10.1038/35012228
|
| [9] |
Gong G C, Lee-Chen Y L, Liu K K. 1996. Chemical hydrography and chlorophyll a distribution in the East China Sea in summer: implications in nutrient dynamics. Continental Shelf Research, 16: 1561–1590. doi: 10.1016/0278-4343(96)00005-2
|
| [10] |
Grasshoff K, Kremling K, Ehrhardt M. 1999. Methods of Seawater Analysis. 3th ed. Chichester: John Wiley & Sons
|
| [11] |
Guo Shujin, Feng Yuanyuan, Wang Lei, et al. 2014. Seasonal variation in the phytoplankton community of a continental-shelf sea: the East China Sea. Marine Ecology Progress Series, 516: 103–126. doi: 10.3354/meps10952
|
| [12] |
Hashihama F, Horimoto N, Kanda J. 2008. Temporal variation in phytoplankton composition related to water mass properties in the central part of Sagami Bay. Journal of Oceanography, 64: 23–37. doi: 10.1007/s10872-008-0002-8
|
| [13] |
Hayakawa M, Suzuki K, Saito H, et al. 2008. Differences in cell viabilities of phytoplankton between spring and late summer in the northwest Pacific Ocean. Journal of Experimental Marine Biology and Ecology, 360: 63–70. doi: 10.1016/j.jembe.2008.03.008
|
| [14] |
Hu Dunxin, Wu Lixin, Cai Wenju, et al. 2015. Pacific western boundary currents and their roles in climate. Nature, 522: 299–308. doi: 10.1038/nature14504
|
| [15] |
Huang Bangqin, Ou Liujian, Hong Huasheng, et al. 2005. Bioavailability of dissolved organic phosphorus compounds to typical harmful dinoflagellate Prorocentrum donghaiense Lu. Marine Pollution Bulletin, 51: 838–844. doi: 10.1016/j.marpolbul.2005.02.035
|
| [16] |
Ito S, Matsuo Y, Yokouchi K, et al. 2000. Cross frontal flow associated with meanders of the Kuroshio Extension and distribution of chlorophyll-a: Observational results from the Wakataka-Maru cruise in May 1997. Bulletin of Tohoku National Fisheries Research Institute (in Japanese), 63: 125–134
|
| [17] |
Jensen K R. 2006. Biogeography of the Sacoglossa (Mollusca, Opisthobranchia). Bonner Zoologische Beiträge, 55: 255–281
|
| [18] |
Justic D, Rabalais N N, Turner R E. 1995. Stioichiometry nutrient balance and origin of coastal eutrophication. Marine Pollution Bulletin, 30: 41–46. doi: 10.1016/0025-326X(94)00105-I
|
| [19] |
Kasai H, Saito H, Yoshimori A, et al. 1997. Variability in timing and magnitude of spring bloom in the Oyashio region, the western subarctic Pacific off Hokkaido, Japan. Fisheries Oceanography, 6(2): 118–129. doi: 10.1046/j.1365-2419.1997.00034.x
|
| [20] |
Kono T, Sato M. 2010. A mixing analysis of surface water in the Oyashio region: its implications and application to variations of the spring bloom. Deep-Sea Research II, 57(17): 1595–1607
|
| [21] |
Lee R E. 2008. Phycology. 4th ed. Cambridge: Cambridge University Press
|
| [22] |
Lee-Chen Y L. 2000. Comparisons of primary productivity and phytoplankton size structure in the marginal regions of southern East China Sea. Continental Shelf Research, 20: 437–458. doi: 10.1016/S0278-4343(99)00080-1
|
| [23] |
Liu Hongbin, Suzuki K, Saito H. 2004. Community structure and dynamics of phytoplankton in the western subarctic Pacific Ocean: a synthesis. Journal of Oceanography, 60(1): 119–137. doi: 10.1023/b:joce.0000038322.7964
|
| [24] |
Marumo R, Asaoka O, Karoji K. 1961. On the distribution of Eucampia zoodiacus Ehrenberg with reference to hydrographic conditions. Journal of Oceanography, 17: 45–47
|
| [25] |
Nakata K. 1988. Alimentary tract contents and feeding condition of ocean-caught post larval Japanese sardinem, Sardinops melanostictus. Bulletin of Tokai Regional Fisheries Research Laboratory, 126: 11–24
|
| [26] |
Nakata K, Hidaka K. 2003. Decadal-scale variability in the Kuroshio marine ecosystem in winter. Fisheries Oceanography, 12(4/5): 234–244
|
| [27] |
Nakata K, Zenitani H, Inagake D. 1995. Difference in food availability for Japanese sardine larvae between the frontal region and the waters on the offshore side of the Kuroshio. Fisheries Oceanography, 4: 68–79. doi: 10.1111/j.1365-2419.1995.tb00061.x
|
| [28] |
Nakayama Y, Kuma K, Fujita S, et al. 2010. Temporal variability and bioavailability of iron and other nutrients during the spring phytoplankton bloom in the Oyashio region. Deep-Sea Research Part II, 57: 1618–1629. doi: 10.1016/j.dsr2.2010.03.006
|
| [29] |
Nishibe Y, Takahashi K, Sato M, et al. 2017. Phytoplankton community structure, as derived from pigment signatures, in the Kuroshio Extension and adjacent regions in winter and spring. Journal of Oceanography, 73: 463–478. doi: 10.1007/s10872-017-0415-3
|
| [30] |
Nishibe Y, Takahashi K, Shiozake T, et al. 2015. Size-fractionated primary production in the Kuroshio extension and adjacent regions in spring. Journal of Oceanography, 71: 27–40. doi: 10.1007/s10872-014-0258-0
|
| [31] |
Okamoto S, Hirawake T, Saitoh S I. 2010. Interannual variability in the magnitude and timing of the spring bloom in the Oyashio region. Deep-Sea Research Ⅱ, 57: 1608–1617. doi: 10.1016/j.dsr2.2010.03.005
|
| [32] |
Redfield A C, Ketchum B H, Richards F. 1963. The influence of organisms on the composition of seawater. In: Hill M N, ed. The Sea. Vol. 2. New York: John Wiley, 26–77
|
| [33] |
Richardson A J. 2008. In hot water: zooplankton and climate change. ICES Journal of Marine Science, 65: 279–295. doi: 10.1093/icesjms/fsn028
|
| [34] |
Round F E, Crawford R M, Mann D G. 1990. The Diatoms: Biology and Morphology of the Genera. Cambridge, UK: Cambridge University Press, 1–747
|
| [35] |
Runge J A. 1980. Effects of hunger and season on the feeding behavior of Calanus pacificus. Limnology and Oceanography, 25: 134–145. doi: 10.4319/lo.1980.25.1.0134
|
| [36] |
Saito H, Tsuda A, Kasai H. 2002. Nutrient and plankton dynamics in the Oyashio region of the western subarctic Pacific Ocean. Deep-Sea ResearchⅡ, 49: 5463–5486
|
| [37] |
Schlundt C, Tegtmeier S, Lennartz S T, et al. 2017. Oxygenated volatile organic carbon in the western Pacific convective center: ocean cycling, air-sea gas exchange and atmospheric transport. Atmospheric Chemistry and Physics, 17: 0837–10854
|
| [38] |
Shen Zhiliang, Zhou Shuqing, Pei Shaofeng. 2008. Transfer and transport of phosphorus and silica in the turbidity maximum zone of the Changjiang estuary. Estuarine Coastal and Shelf Science, 78: 481–492. doi: 10.1016/j.ecss.2008.01.010
|
| [39] |
Siswanto E, Honda M C, Sasai Y, et al. 2016. Meridional and seasonal footprints of the Pacific Decadal Oscillation on phytoplankton biomass in the northwestern Pacific Ocean. Journal of Oceanography, 72: 465–477. doi: 10.1007/s10872-016-0367-z
|
| [40] |
Smayda T J. 1997. Harmful algal blooms: their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnology and Oceanography, 42: 1137–1153. doi: 10.4319/lo.1997.42.5_part_2.1137
|
| [41] |
Stoecker D K. 1999. Mixotrophy among dinoflagellates. Journal Eukaryot Microbiol, 46: 397–401. doi: 10.1111/j.1550-7408.1999.tb04619.x
|
| [42] |
Sun Jun, Liu Dongyan. 2002. The preliminary notion on nomenclature of common phytoplankton in China seas waters. Oceanologia et Limnologia Sinica (in Chinese), 33(3): 271–286
|
| [43] |
Sun Jun, Liu Dongyan. 2004. The application of diversity indices in marine phytoplankton studies. Acta Oceanologica Sinica, 26: 62–75
|
| [44] |
Takahashi M, Nishida H, Yatsu A, et al. 2008. Year-class strength rates after metamorphosis of Japanese sardine (Sardinops melanosticus) in the western North Pacific Ocean during 1996–2003. Canadian Journal of Fisheries and Aquatic Sciences, 65: 1425–1434. doi: 10.1139/F08-063
|
| [45] |
Takahashi T, Sutherland S C, Sweeney C, et al. 2002. Global sea-air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects. Deep-Sea Research Part II, 49: 1601–1622. doi: 10.1016/S0967-0645(02)00003-6
|
| [46] |
Taniguchi A. 1999. Differences in the structure of the lower trophic levels of pelagic ecosystems in the eastern and western subarctic Pacific. Progress in Oceanography, 43: 289–315. doi: 10.1016/S0079-6611(99)00011-7
|
| [47] |
Taniguchi A, Kawamura T. 1972. Primary production in the Oyashio region with special reference to the subsurface chlorophyll maximum layer and phytoplankton-zooplankton relationships. In: Takenouti A, ed. Biological Oceanography of the Northern North Pacific Ocean. Tokyo: Idemitsu Shoten, 419–431
|
| [48] |
Tittensor D P, Mora C, Jetz W, et al. 2010. Global patterns and predictors of marine biodiversity across taxa. Nature, 466(7310): 1098–1101. doi: 10.1038/nature09329
|
| [49] |
Tomas C R. 1997. Identifying Marine Phytoplankton. San Diego: Academic Press, 1–858
|
| [50] |
Utermöhl H. 1958. Zur vervollkommung der quantitativen phytoplankton-methodik. Mitteilungen der Internationale Vereinigung für Theoretische und Angewandte Limnologie, 9: 1–38
|
| [51] |
Watanabe Y. 2007. Latitudinal variation in the recruitment dynamics of the small pelagic fishes in the western North Pacific. Journal of Sea Research, 58: 46–58. doi: 10.1016/j.seares.2007.02.002
|
| [52] |
Watanabe Y, Kurita Y, Noto M, et al. 2003. Growth and survival processes of Pacific saury Cololabis saira in the Kuroshio-Oyashio Transition waters. Journal of Oceanography, 59: 403–414. doi: 10.1023/A:1025532430674
|
| [53] |
Yamamoto T, Nishizawa S, Taniguchi A. 1988. Formation and retention mechanisms of phytoplankton peak abundance in the Kuroshio front. Journal of Plankton Research, 10: 1113–1130. doi: 10.1093/plankt/10.6.1113
|
| [54] |
Yasuda I. 2003. Hydrographic structure and variability in the Kuroshio-Oyashio transition area. Journal of Oceanography, 59: 389–402. doi: 10.1023/A:1025580313836
|
| [55] |
Yasuda I, Okuda K, Hirai M. 1992. Evolution of a Kuroshio warm-core ring—variability of the hydrographic structure. Deep-Sea Research, 39(Suppl 1): S131–S161
|
| [56] |
Yin Kedong, Qian Peiyuan, Chen J C, et al. 2000. Dynamics of nutrients and phytoplankton biomass in the Pearl River estuary and adjacent waters of Hong Kong during summer: preliminary evidence for phosphorus and silicon limitation. Marine Ecology Progress Series, 194: 295–305. doi: 10.3354/meps194295
|
Figures(5) / Tables(3)

Supported by:
Beijing Renhe Information Technology Co. Ltd

Yongxu Li, Xudong Lai, Mingwei Wang. Semisupervised heterogeneous ensemble for ship target discrimination in synthetic aperture radar images[J]. Acta Oceanologica Sinica, 2022, 41(7): 180-192. doi: 10.1007/s13131-021-1980-2

| Characteristics | P. commixtus | P. chenii sp. nov. | |
| Carapace | left asymmetric gland | near postero-dorsal margin, moved anteriorly | on postero-dorsal corner |
| right asymmetric gland | on right postero-ventral corner | none | |
| right lateral gland | constituting a compound gland with right asymmetric gland | only on right postero-ventral corner | |
| Frontal organ | stem and capitulum | fused | separated |
| spines | small disto-ventral and mid-ventral spines | small disto-dorsal and ventral spines | |
| First antenna | endopod 2 | with one long dorsal plumose seta | with one small dorsal plumose seta |
| sensory setae | a- to d-setae analogic, long columnar, thin walled and bare, with long end joint; e-seta bare, with short end joint | a- and c-setae analogic, long columnar, thin walled and bare with long end joint; b- and d- setae analogic, equilong and bare without end joint, slightly longer than a- and c-setae; e- seta spinose, without short end joint | |
| Second antenna | endopod | c-, d-, e-setae absent; b-seta bare; one small oval hump with central concave on mid-ventral margin, instead of processus mamillaris | e-setae present; b-seta spinose; processus mamillaris normal. |
| exopod | exopod 1 and 2 fused, exopod 2-4 bare, exopod 8 and 9 separated; terminal plumose seta with single tip | exopod 1 and 2 separated, exopod 2-4 with plumose setae, exopod 8 and 9 fused | |
| Mandible | coxale teeth side | with eight distal teeth | with six distal teeth |
| tooth endites | with four tooth plates | with three tooth plates | |
| endopod | endopod 1 with one ventral seta, and one dorsal seta; endopod 2 with one disto-ventral seta; terminal segment with seven spinose setae | endopod 1 with one ventral seta, without dorsal seta; endopod 2 with two disto-ventral setae; terminal segment with six setae, ventral three bare | |
| Maxilla | endopod | endopod 1 with six disto-anterior setae, one disto-medial seta, none medial seta, and two disto-posterior setae; mid seta of terminal segment bare | endopod 1 with five disto-anterior setae, one disto-medial seta, two medial setae, and three disto-posterior setae; mid seta of terminal segment spinose |
| endites | endite I with eleven plumose setae, endite II with ten plumose setae, endite III with eight plumose setae | endite I with five plumose setae; endite II with one bare and three plumose setae; endite III with ten bare setae | |
| Fifth limb | basale | proximal-ventral group of setae with four setae | proximal-ventral group of setae with seven setae |
| endopod | endopod 1 with one ventral seta | endopod 1 with two ventral setae | |
| endites | endite I with one seta; endite II with two setae; endite III with one proximal seta, three sharp and two blunt distal setae | endite I with five setae; endite II with one seta; endite III with none proximal seta, four sharp and two blunt distal setae | |
| Sixth limb | basale | disto-dorsal seta (vesting of exopod) small and bare; three small ventral setae. | disto-dorsal seta (vesting of exopod) strong and spinose; seven ventral plumose setae |
| endopod | endopod 1 with one ventral seta; endopod 2 with one ventral seta | endopod 1 bare; endopod 2 without ventral seta | |
| Furca | unpaired seta | no | yes |
DownLoad:
CSV
 DownLoad:
DownLoad: