Figure
1.
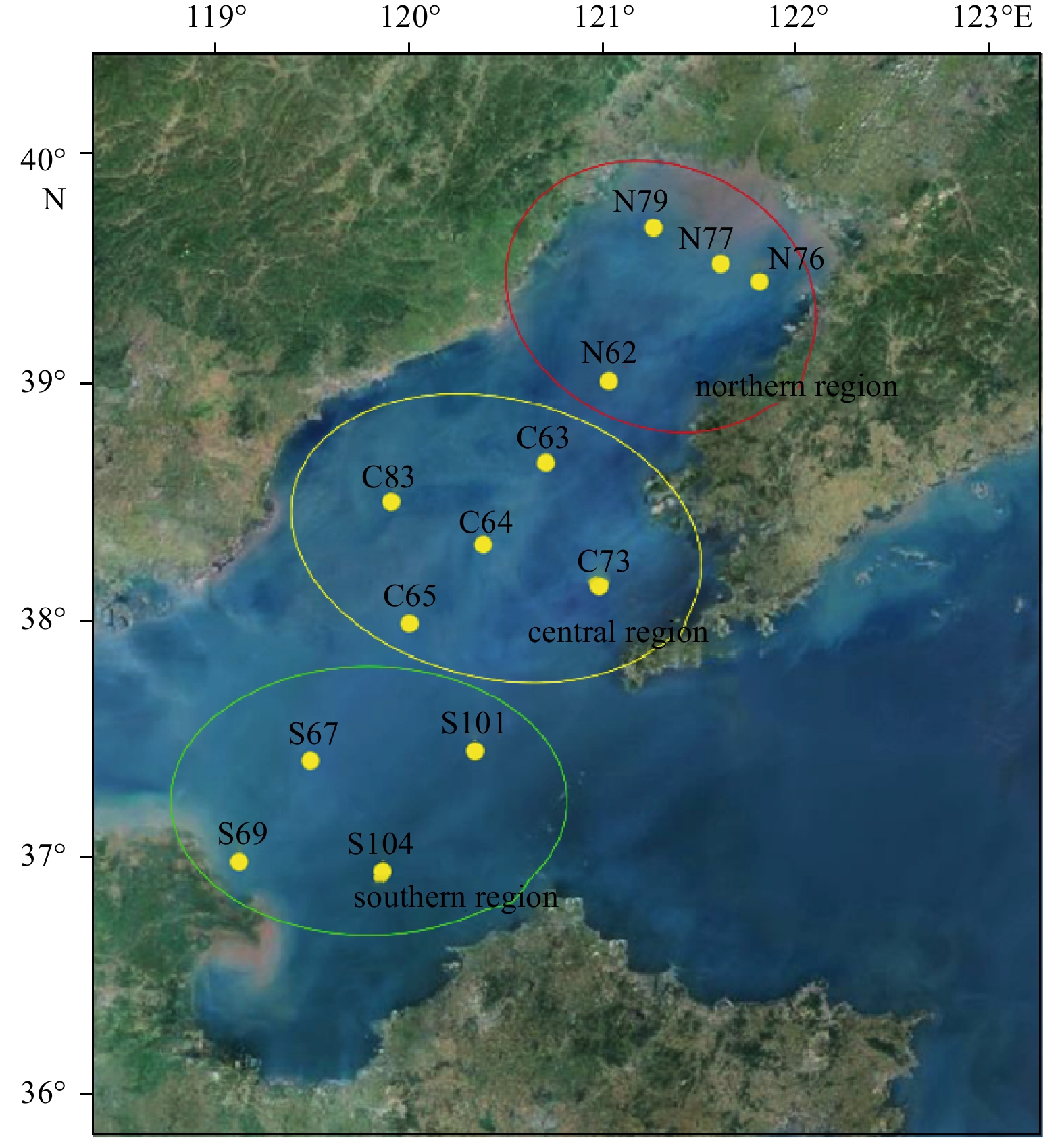
Map of sample collection stations in the Bohai Sea, China.
| Citation: | Quanrui Chen, Jingfeng Fan, Jie Su, Hongxia Ming, Zhihao Sun, Mengfei Li, Xiaohui Zhao, Yantao Wang, Yingxue Zhang, Huizhen Zhang, Yuan Jin, Xiaowan Ma, Bin Wang. Spatial distribution characteristics of bacterial community structure and gene abundance in sediments of the Bohai Sea[J]. Acta Oceanologica Sinica, 2020, 39(2): 69-78. doi: 10.1007/s13131-020-1554-8

|
Coastal ecosystems are extremely important, both in terms of their ecological significance and their economic value to humans (Lv et al., 2016). Sediments comprise an important part of this ecosystem and are subject to interactions between geological, hydrodynamic, biological and chemical factors (Köster and Meyer-Reil, 2001). Offshore sediments are both a source and sink for a large number of pollutants, including nutrients, heavy metals, and petroleum or pesticide residues (Lofrano et al., 2017). Microorganisms participate in various biogeochemical cycles in sediments such as carbon fixation (Jing et al., 2018), methanogens (Tian et al., 2014), nitrification and denitrification (Fan et al., 2011), and biotransformation for a variety of organic pollutants (Wada et al., 2012). Microbial communities are known to be impacted by various ecological factors (Wang et al., 2018), with human activities also producing changes in sediment microbial communities (Yang et al., 2000). Studies of microbial diversity in marine sediments are critical to understanding the evolutionary and ecological processes that affect global microbial diversity, and the response of marine ecosystems to environmental change.
As a semi-open sea in northern China, a large number of population centers have become established on the shores of the Bohai Sea. The GDP of surrounding cities has grown rapidly with the development of industry and agriculture. Recent studies have reported that the Bohai Bay is polluted by heavy metals to varying degrees (Zhou et al., 2017; Li et al., 2017). Heavy metals have also been reported in the nutrient salts of the Bohai Sea. Microbial communities are often considered to have better adaptability and tolerance because they have metabolic pathways that can utilize various resources (Dai et al., 2016; Marshall et al., 2018). Changes in the microbial community under the influence of long-term ecological factors and the potential for selective exposure to different heavy metal contaminants in the environment remains unclear. In this study, the Bohai Sea was selected as the research area. Real-time quantitative PCR and high-throughput sequencing of bacterial 16S rRNA genes were used to study the community abundance and structure of bacteria in the sediments, and the influence of ecological factors was explored. The interrelationship between bacterial communities and ecosystems in sediments aids our understanding of the geographical distribution patterns of marine microbes and the niche differentiation of various microbial communities.
The collection of experimental samples was carried out in late May 2017. A total of 13 stations (Fig. 1), were located throughout the Liaodong Bay, the Huanghe (Yellow) River Estuary, the periphery of the Laizhou Bay and the central part of the Bohai Sea. The average depth of the sampling stations was 20 m. The sediments at 0–10 cm of the surface layer were collected using a grab sampler. Three replicate samples were collected at each station. The three samples from each station were mixed together and dispensed into two labelled sterile valve bags, shipped back to the laboratory and stored at –20°C for future analysis.
Station depth was measured at each sampling site and the particle size of the sediment was measured using a LS13320 laser diffraction particle size analyzer (Beckman Coulter, USA). Total organic carbon (TOC), total phosphorus (TP) and total nitrogen (TN) in the sediment, as well as heavy metals including cadmium (Cd), chromium (Cr), copper (Cu), lead (Pb), zinc (Zn), mercury (Hg), arsenic (As), were determined following “Marine Monitoring Specification Part 5: Sediment Analysis GB 17378.5–2007”(General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China and Standardization Administration of China, 2008). Determination of nutrient salts the nitrates and nitrous acid salt and ammonia nitrogen content in the sediments were determined by the method of “Measurement of Soil Ammonium, Nitrite and Nitrate Chloride Solution Extraction-spectrophotometry HJ 634–2012” (Ministry of Environmental Protection of the People’s Republic of China, 2012).
Total bacterial DNA was extracted from sediment samples using the DNeasy PowerSoil Kit (Qiagen, Germany). After the sediment was mixed, 0.35 g of each sample was taken for total DNA extraction, according to the kit extraction instructions. This was carried out in triplicate. After extraction, the quality of the DNA was tested by 1.5% agarose gel electrophoresis. Three DNA samples were then combined into one and stored at –80°C.
The extracted total DNA template was subjected to two-step PCR amplification using a V4-V5 region-specific primer of the bacterial 16S rRNA gene to construct a 16S rRNA gene library (Liu et al., 2008). The first step uses specific primer amplification: 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 926R (5′-CCGTCAATTCMTTTGAGTTT-3′). PCR amplification conditions: 5× buffer, 10 μL; dNTP (10 mmol/L), 1 μL; Phusion ultra-fidelity DNA polymerase 1 U; 1 μL of each primer (10 μmol/L); 5 μL of total DNA template; and finally supplemented with 50 μL ddH2O. The initial PCR procedures were: 94°C for 2 min; follow by 25 cycles of denaturation at 94°C for 30 s; 56°C annealing for 30 s; 72°C for 30 s; and final extension at 72°C for 5 min. The first PCR product was electrophoresed using a 2% detection agarose gel and recovered using the AxyPrep DNA Gel Extraction Kit (Axygen, USA). The second PCR amplification was then performed, to add the linker barcode required for sequencing the Illumina high throughput sequencing platform to the target fragment. After the product was purified, the quantitative control was carried out. Finally, the 16S rRNA gene v4-v5 region was sequenced by TinyGene Bio-Tech (Shanghai) Co., Ltd. through the Illumina MiSeq platform using 2×300 bp end pairing sequencing.
Using the sequencing data, the sample was first distinguished according to the barcode, and the optimized sequence was obtained by splicing, quality control and filtering. The optimized sequence was then clustered using Mothur (version 1.39.5) and USEARCH programs, these sequences were clustered to OTUs (operational taxonomic units) at 97% sequence identity (furthest neighbor method), and then the classification level of OTU in each sample was classified according to the species level. Mothur and R language were used to plot the dilution curve and abundance level map of OTUs, and multi-sample similarity analysis, principal component analysis and alpha diversity were completed by OTU classification level results. Environmental factors and RDA analysis between heavy metals and samples were performed using CANOCO 5.0 software, and SPSS 23.0 for correlation analysis between various factors.
The heavy metal pollution assessment was calculated by the geoaccumulation index (Igeo) method (Muller, 1969), and the calculation equation is as follows:
| $${I_{{\rm{geo}}}} = {\log _2}\left[ {\frac{{\rho_n}}{{k \times B_n}}} \right],$$ | (1) |
where ρn represents the concentration of metal (n) in the sample, Bn is the background value of the element in the sediment, k is used to correct the background value of the region, and the sediment environment is usually taken as 1.5 (Müller and Förstner, 1976).
The original sequence of the bacterial 16S rRNA gene obtained by high-throughput sequencing in this study has been registered in the NCBI database, and the gene sequence accession number is SRR6323824 to SRR6323836 (13 samples).
The environmental factors of sediment samples from 13 stations in the Bohai Sea are shown in Table 1. Nitrate content in the sediments of the Bohai Sea region was between 138–513 μg/g and the ammonia nitrogen content was 72–197 μg/g. Nitrate and ammonia nitrogen were highest in the southern region, with lowest values recorded in the central region. Nitrous acid salt content ranged from 27.6–103 μg/g in the central region. TP was higher near sampling Stas S69 and S104. Overall TP values were highest in the southern region, followed by the northern region, and were lowest in the central region. Highest values for TN, TOC and grid size were recorded at sampling Sta. C83 near Qinhuangdao. The sampling stations ranged in depth from 10 to 34 m, and the central region was significantly deeper than other areas. According to the on-site measurement data, there was no significant difference in salinity and pH between stations, so it was not included in the analysis.
| Station | NO3–/μg·g–1 | NO2–/μg·g–1 | NH4+/μg·g–1 | TOC/% | TP/μg·g–1 | TN/μg·g–1 | depth/m | Grid size (Mz, Φ) |
| S67 | 421 | 29.9 | 153 | 0.66 | 143 | 130 | 23 | 6.63 |
| S69 | 513 | 27.6 | 173 | 0.43 | 386 | 122 | 10 | 5.76 |
| S101 | 482 | 59 | 140 | 0.36 | 174 | 93.6 | 27 | 5.64 |
| S104 | 256 | 29.9 | 121 | 0.53 | 348 | 135 | 17.5 | 6.11 |
| C63 | 217 | 68.9 | 96.2 | 0.49 | 255 | 97.3 | 28 | 6.22 |
| C64 | 149 | 85.3 | 101 | 0.24 | 181 | 91.6 | 19.8 | 4.99 |
| C65 | 228 | 70.7 | 104 | 0.39 | 208 | 93.6 | 19 | 4.64 |
| C73 | 183 | 103 | 68.3 | 0.62 | 221 | 145 | 34 | 6.21 |
| C83 | 330 | 83.4 | 93.3 | 0.92 | 269 | 157 | 23 | 7.37 |
| N62 | 185 | 75 | 84.9 | 0.36 | 212 | 95.8 | 26 | 5.43 |
| N76 | 350 | 75.4 | 112 | 0.84 | 290 | 114 | 13 | 6.3 |
| N77 | 256 | 67.4 | 72 | 0.53 | 219 | 100 | 12.7 | 5.81 |
| N79 | 138 | 58.5 | 197 | 0.75 | 245 | 111 | 12.5 | 6.52 |
 DownLoad:
CSV
DownLoad:
CSV
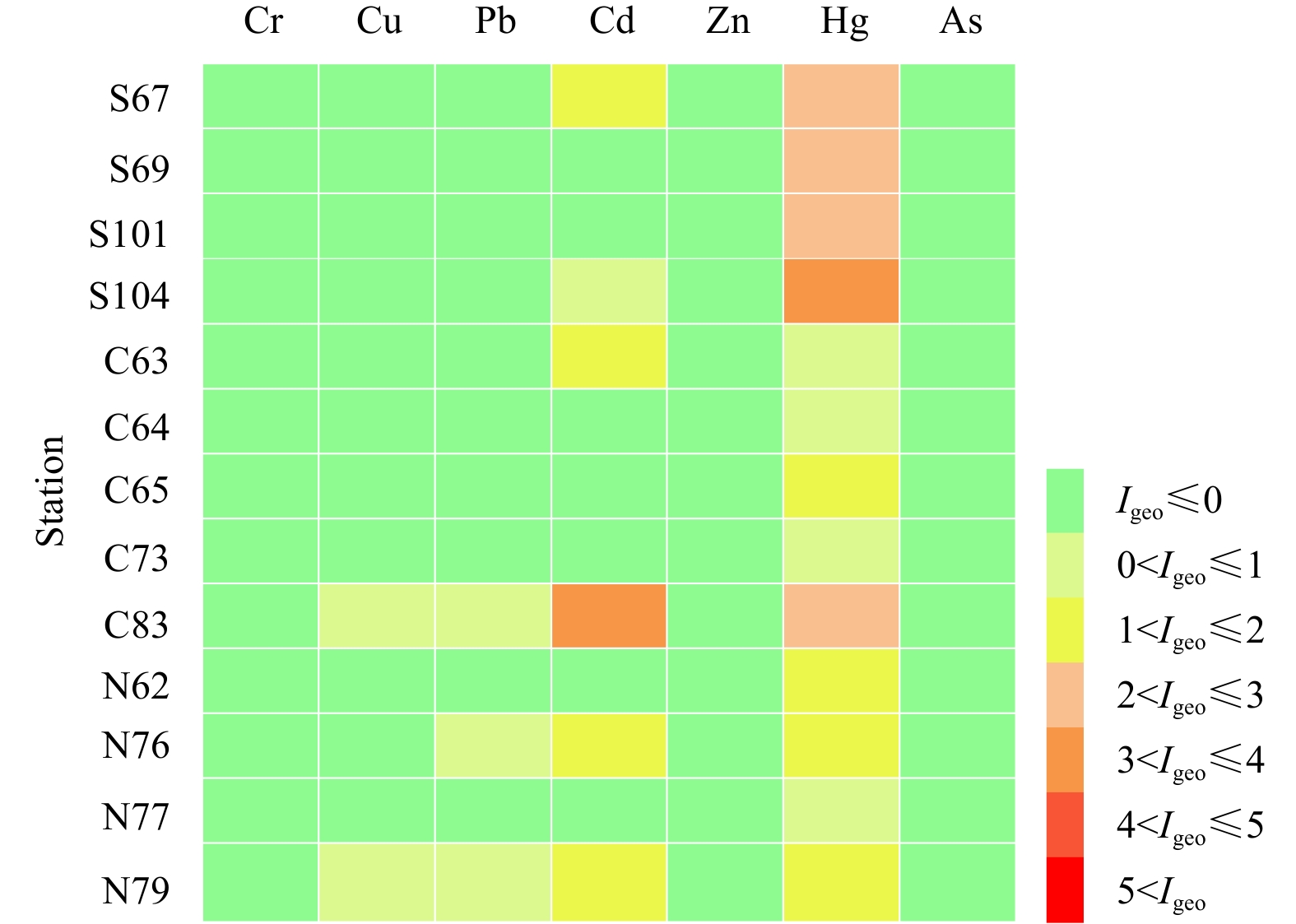
According to the results of the geoaccumulation index method in the sediment (Fig. 2), heavy metals such as Cr, Zn and As in the sediment of the 13 sampling sites in the Bohai Sea could be classified as unpolluted (Igeo≤0). Cu was present at low pollution levels at sampling Stas N79 and C83 (0<Igeo≤1), while Pb was also recorded as slightly polluting at three sampling stations (0<Igeo≤1). Six sampling stations were found to be moderately polluted by Cd. In particular, Sta. N79 near the coast of Jinzhou in the Liaodong Bay was highly polluted. Hg was detected at all sampling stations. Stations S104 and S101 in the Laizhou Bay area had the highest concentrations of Hg. The degree of Hg pollution was highest in the southern region, followed by the northern region, and was lowest in the central region. Stations N77, C64 and C73 had low levels of heavy metal pollution, with only mild contamination of Hg. Sampling Sta. C83 was affected by a variety of heavy metal pollutants.

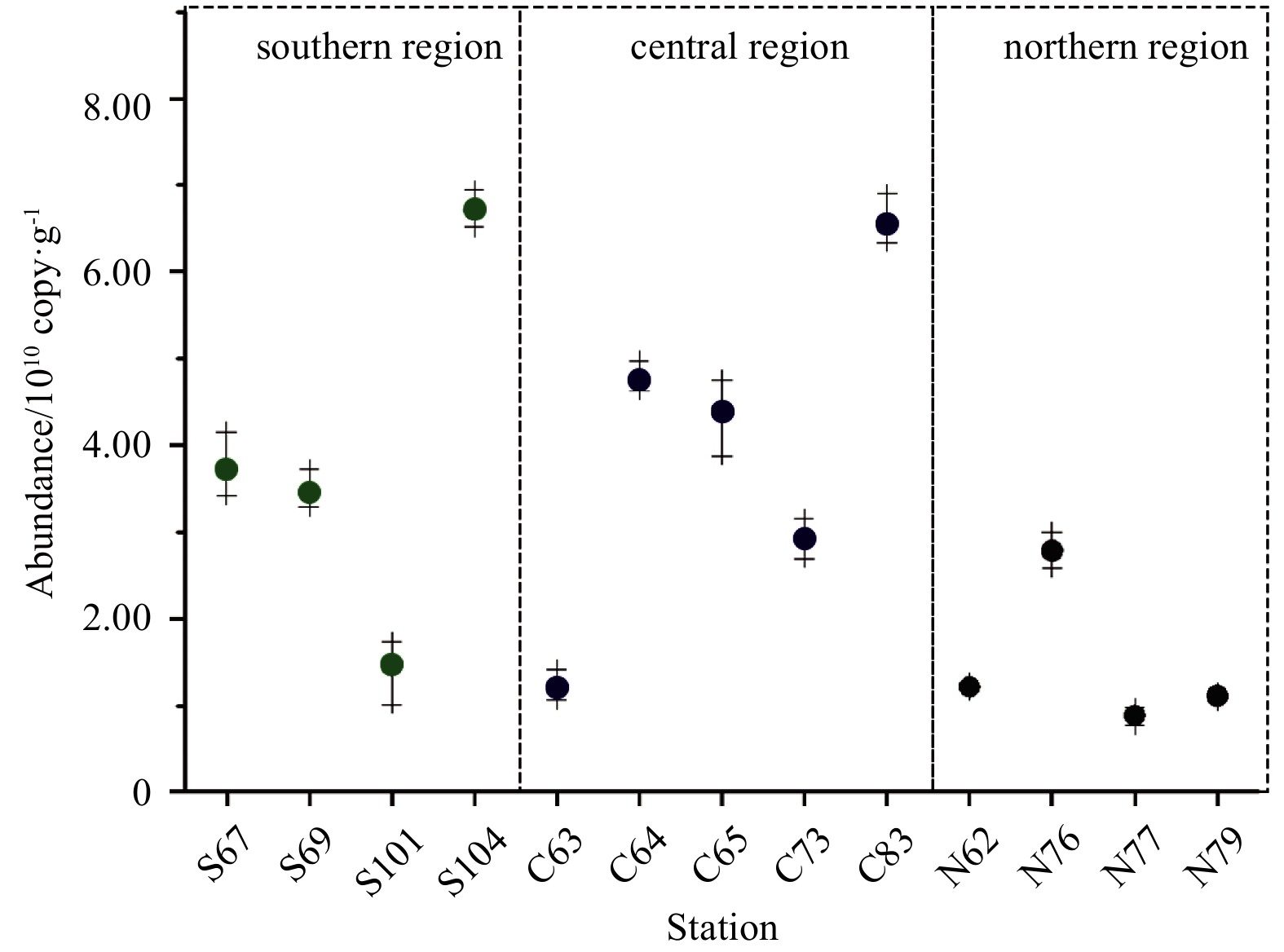
The gene abundance of the bacterial 16S rRNA gene V4–V5 region from the 13 sampling stations was determined by real-time PCR (qPCR) as shown in Fig. 3. The gene abundance of the 16S rRNA gene V4–V5 region in the sediment at sampling stations in the Bohai Sea ranged from 8.83×109 to 6.71×1010 copy/g, with an average abundance of 3.17×1010 copy/g. The gene abundance in the northern region was found to be lower than that in the southern and central regions. Sampling Sta. C63 in the central region, located close to the northern region, had the lowest gene abundance. The sediments of sampling Stas C83 and S104 were rich in bacterial 16S abundance. The only ecological factor that showed a significant correlation with the gene abundance at each sampling station was TN (P<0.05).

Samples from the 13 sampling sites in the Bohai area were sequenced by high-throughput sequencing of the bacterial 16S rRNA gene V4–V5 region of the extracted total DNA. As shown in Table 2, the number of optimized sequences obtained was 22 514–32 385. All sequences were clustered by 97% similarity to obtain 3 041 different OTUs. The number of OTUs at each sampling station ranged from 1 585 to 2 182, and the highest value was recorded at Sta. S67. This indicated that the sediments at this site had rich bacterial diversity, while Sta. S69 near the Huanghe River Estuary had the lowest number of OTUs (1 585). A maximum of 1 937 OTUs was recorded in the southern region, which was significantly higher than that in the northern region. The coverage of all samples was above 97.7%, indicating that the sample library coverage was high, and most of the bacterial 16S rRNA gene sequences were detected at all sites. The sequencing results were well representative and reliable. The Chao1 index showed that Sta. S67 had the highest total number of species, while the Shannon index showed that Sta. C83 had the highest bacterial diversity. The Shannon index of all stations was between 5.59 and 6.24. The Shannon index value was highest for the southern region, followed by the northern region, with the lowest value obtained for the central region, meaning that the relative average bacterial diversity in the central region was lower than that in the southern and northern regions. The Chao1 index showed a significant correlation with the Shannon index between stations (P<0.05).
| Station | Reads | OTU | Ace | Chao1 | Shannon | Simpson | Coverage/% |
| S67 | 28 770 | 2 182 | 2 547 | 2 520 | 6.13 | 0.008 2 | 98.2 |
| S69 | 22 563 | 1 585 | 2 029 | 2 079 | 5.83 | 0.009 9 | 97.9 |
| S101 | 32 385 | 1 963 | 2 299 | 2 328 | 5.94 | 0.008 7 | 98.6 |
| S104 | 22 514 | 2 017 | 2 395 | 2 397 | 6.1 | 0.008 6 | 97.7 |
| C63 | 26 467 | 2 011 | 2 371 | 2 347 | 6.07 | 0.008 8 | 98.2 |
| C64 | 24 761 | 1 812 | 2 245 | 2 208 | 5.59 | 0.016 1 | 97.9 |
| C65 | 25 325 | 1 825 | 2 282 | 2 248 | 5.78 | 0.009 8 | 98.0 |
| C73 | 28 309 | 1 838 | 2 345 | 2 370 | 5.62 | 0.014 7 | 98.1 |
| C83 | 24 577 | 2 117 | 2 473 | 2 505 | 6.24 | 0.007 2 | 98.0 |
| N62 | 24 563 | 1 727 | 2 135 | 2 113 | 5.78 | 0.010 8 | 98.1 |
| N76 | 26 143 | 2 065 | 2 457 | 2 428 | 6.07 | 0.009 2 | 98.0 |
| N77 | 24 527 | 1 892 | 2 325 | 2 279 | 5.92 | 0.011 3 | 97.9 |
| N79 | 26 312 | 1 873 | 2 257 | 2 239 | 5.93 | 0.009 3 | 98.2 |
DownLoad:
CSV
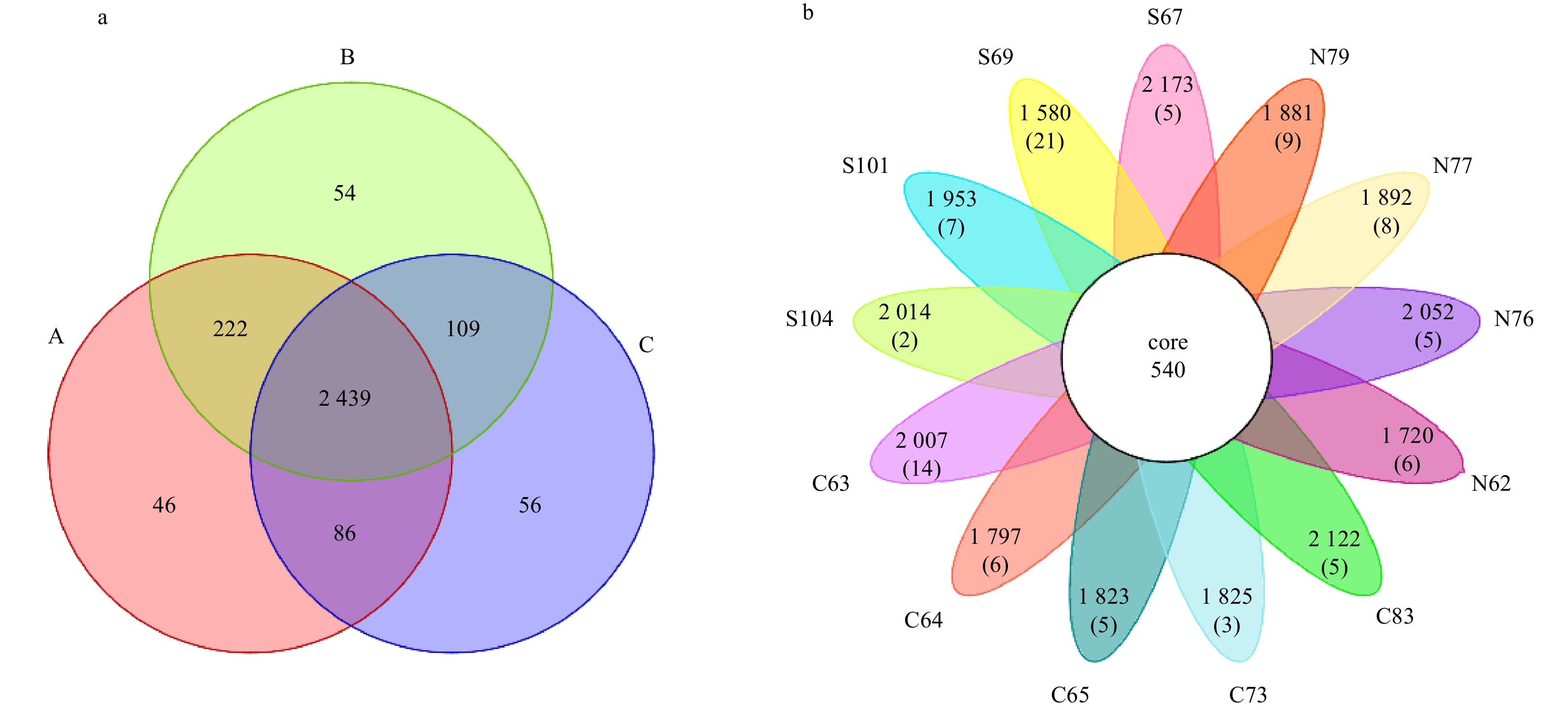
The Venn diagram shows the number of common and unique OTUs in each sediment sample. As shown in Fig. 4a, 2 439 OTUs were common to all three regions. The southern and central regions had 222 common OTUs, which is higher than the overlap of the northern and central and northern and southern regions. There were 56 unique OTUs in the northern region, indicating that the northern region had relatively more endemic microbial species than the central or southern regions. The petal diagram in Fig. 4b shows the number of OTUs shared between the 13 sampling stations. The core microbiome analysis found 100% of the OTUs covering the 13 samples. The number of core OTUs was 540. Station S69, near the mouth of the Huanghe River in the southern region, had the highest number of unique OTUs (21), followed by Sta. C63 in the central region, which had 14 unique OTUs.

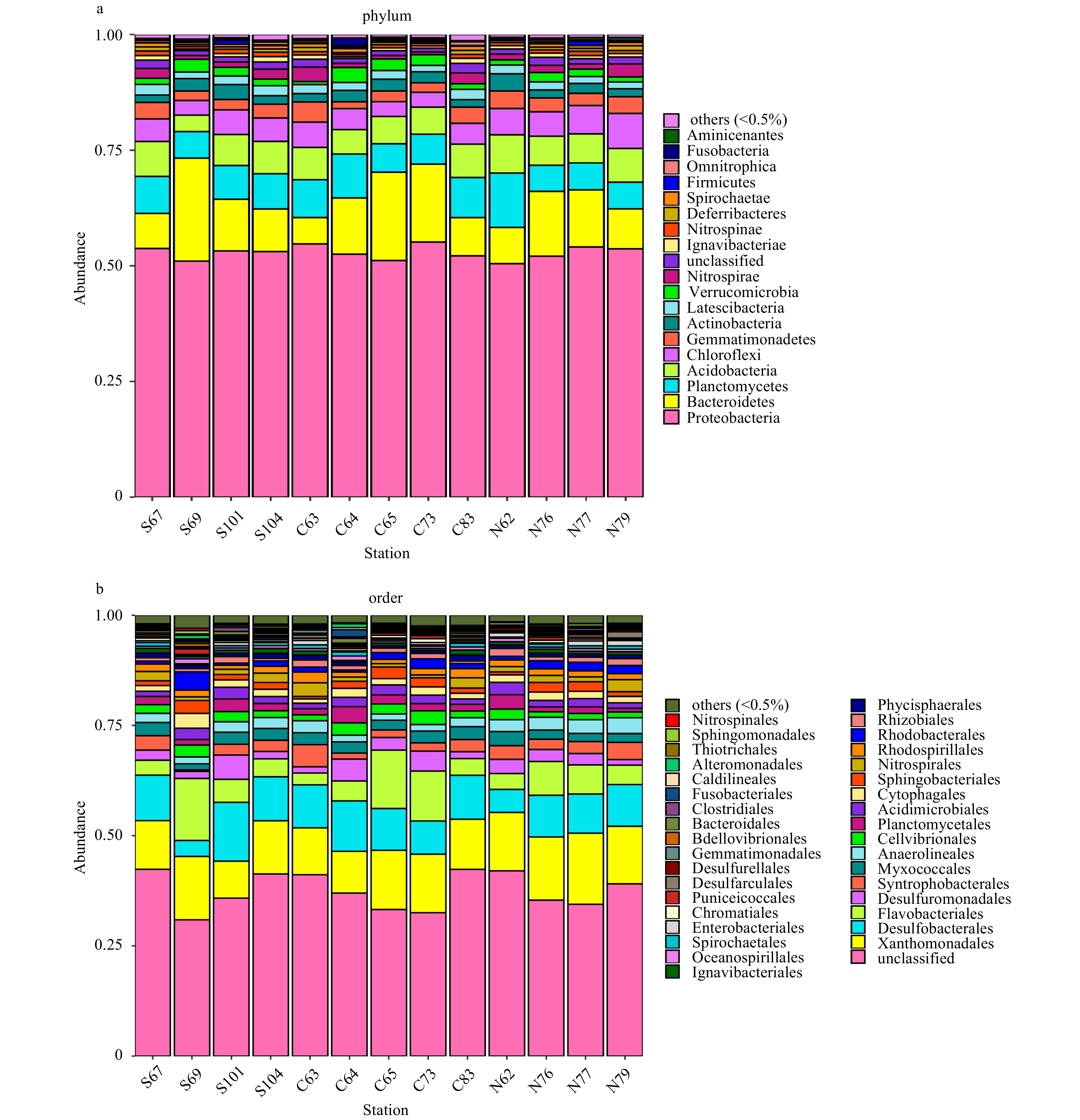
The OTUs of the bacterial communities in the sediments of the 13 sampling stations were clustered into 36 phyla, 49 classes, 77 orders, 109 families, 181 genera and 198 species. The main taxonomic results are shown in Fig. 5a, in which Proteobacteria dominated the sediment samples from the Bohai Sea, with an average relative abundance of 52.92%. The average relative abundance of Bacteroidetes was 11.76%. The relative abundance of Planctomycetes was 7.39%, Acidobacteria had a relative abundance of 6.53%, while Chloroflexi had a relative abundance of 4.97%. The remaining bacteria had a relative abundance of less than 4.0%. In the central region, at Sta. S69, Bacteroidetes were most abundant (22.29%), and were significantly more abundant here than other sampling stations. A total of 77 bacterial orders (including unclassified parts) were detected at the 13 sampling stations. The main orders are shown in Fig. 5b. Xanthomonadales were dominant at all sampling stations (average relative abundance of 12.29%), followed by Desulfobacterales (9.23%), Flavobacteriales (6.41%) and Desulfuromonadales (2.83%). Station S69 had the lowest relative abundance of Desulfobacterales (3.59%) and the highest relative abundance of Flavobacteriales (14.07%).

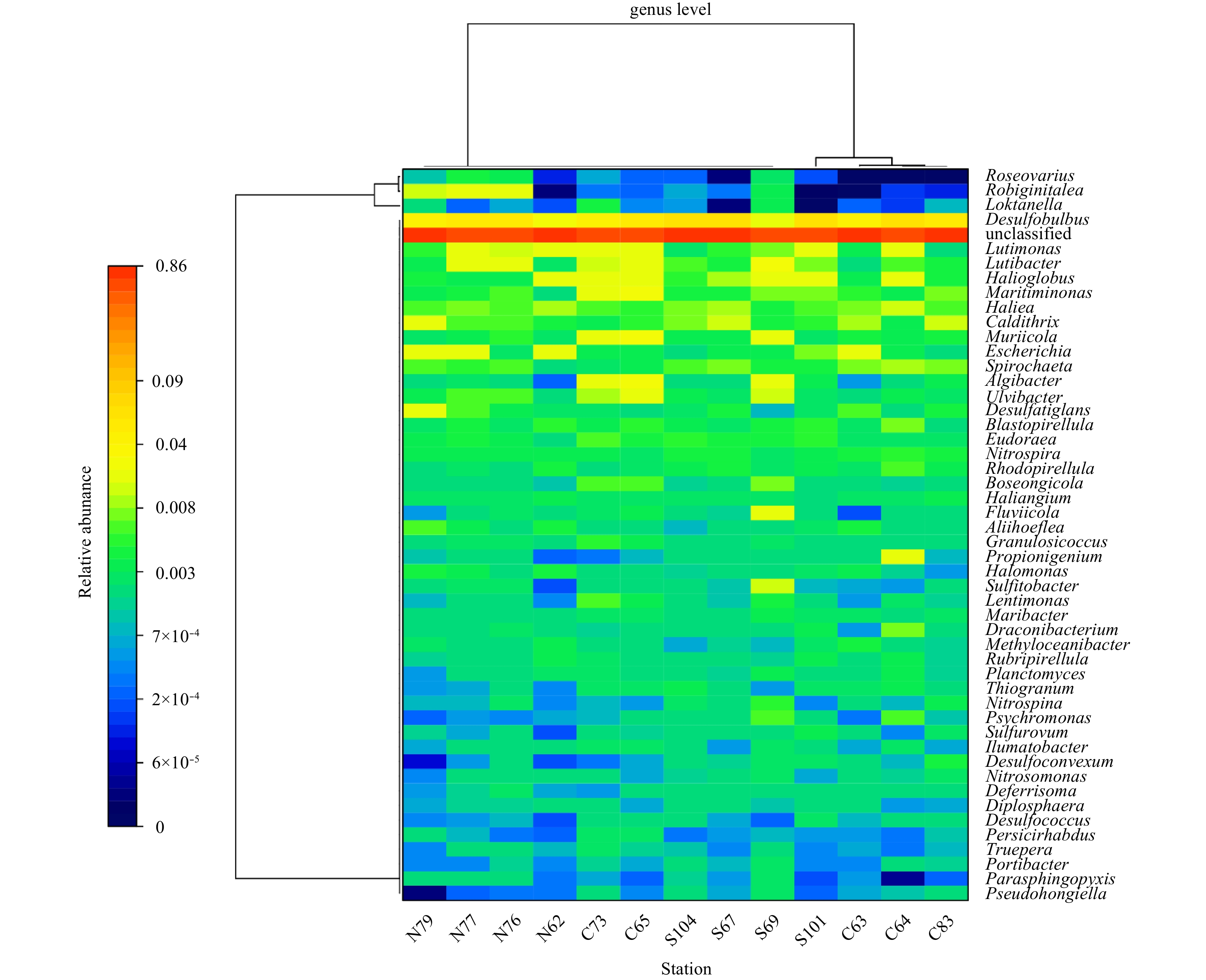
As shown in Fig. 6, at the genus level, the Desulfobulbus of the δ-Proteobacteria was the only genus with a relative abundance higher than 1%. Desulfobulbus had a relative abundance of 4.99%, followed by the genus Lutimonas (0.88%) of the family Flavobacteriia, the genus Halioglobus (0.84%) of the γ-proteobacteria, and the genus Lutibacter of the family Flavobacteriia

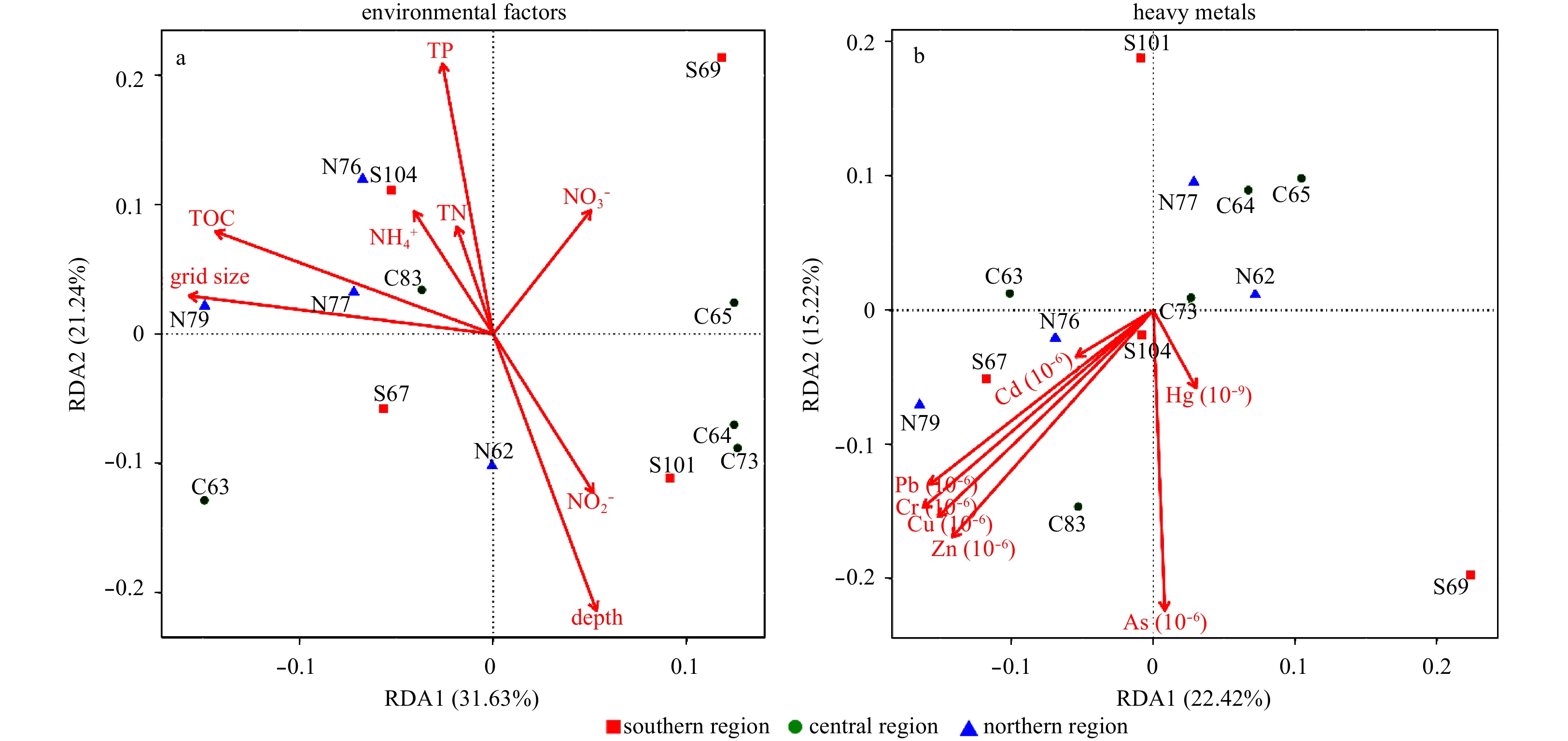
Some of the environmental factors of the sediments in the Bohai Sea region varied greatly and were influenced by bacterial community structure. Redundancy analysis of environmental factors and the bacterial community structure at each sampling station was carried out using CANOCO 5.0 software (Fig. 7a). The degree of interpretation of the sorting axes RDA1 and RDA2 were 31.63% and 21.24%, respectively. The composition of the bacterial community at Sta. S69 was quite different from the other sampling stations, and was mainly affected by the positive correlation of nitrate content. The environmental factors that were positively correlated with the community structure at Stas N76, N77 and N79 were TOC and sediment size, followed by ammonia nitrogen, TN and TP. Stations C62, C64, C73 and S101 had a significant positive correlation with sediment depth and a significant negative correlation with TOC and grid size. For all environmental factors, TOC had the highest significant correlation with bacterial community structure (P=0.027, P<0.05), while there was no significant correlation with ammonium (P=0.738), nitrate (P=0.394) and nitrite (P=0.269) and the bacterial community in the sediments of the Bohai Sea. The results showed that the main factor influencing bacterial community structure in the sediments of the Bohai Sea was the TOC content in the sediment, followed by depth and TP.

Redundancy analysis of heavy metal content and bacterial community structure at each sampling station was also carried out with CANOCO 5.0 software (Fig. 7b). The degree of interpretation of the sorting axes RDA1 and RDA2 were 22.42% and 15.22%, respectively. It can be seen that the sequence of influence of heavy metals on the microbial community structure in the sediment was Zn, Cr, As, Cu, Pb, Hg and Cd. Correlation with bacterial communities showed consistency in the effects of Pb, Cd, Cr, Cu and Zn. There was a positive correlation between the bacterial community at Stas C83 and S104 and heavy metals (P<0.05). The sediments at these two sampling stations were heavily contaminated by heavy metals. The negative correlation between Stas C65, C64 and C73 and most heavy metals were not significantly associated with Hg. The bacterial community structure at Sta. S101 was significantly negatively correlated with As. In the majority of samples the content of arsenic was negatively correlated with bacterial community structure; however, the influence of Hg on community structure was not significant. Although the pollution of Hg and Cd was the most obvious in each sediment samples, the effects of these two heavy metals on the bacterial community structure were not significant, this result may be the stress response of bacterial community structure to heavy metals, and is a long-term accumulation and evolution process, but this conclusion needs to be further confirmed by metagenomics data in the future.
The bacterial community structure in marine sediments drives a variety of biogeochemical cycles. The study of the potential distribution patterns of these microorganisms and the mechanisms by which their community structure changes in the environment has always received much attention (Shade, 2018). The results of the current study showed that a large number of bacterial OTUs (1 585–2 182) were detected in sediments of the Bohai Sea. Diversity was highest in the southern and central regions, with bacterial 16S rRNA gene abundance showing the same trend. In studies of the Liaohe Estuary of the Bohai Sea, using high-throughput sequencing of the bacterial 16S rRNA gene V3-V4 region our laboratory found that the diversity of bacterial communities in the sediments farther from the shore was lower than estuarine, freshwater and mixed areas. A total of 32 phyla were recorded in the previous studies, lower than the 36 phyla in the current study. The diversity of sedimentary bacterial communities in the northern region of the current study coincided with the previous results from the Liaohe Estuary (Zhang et al., 2018). A relatively high number of OTUs (2 590–3 777) has been reported from the sediments of the Liaodong Bay in the Bohai Sea (Zheng et al., 2014). The number of OTUs obtained by 454 pyrosequencing from three sediment deposits in the Bohai Sea ranged from 3 371–5 224 (Liu et al., 2015). The main reason for the difference was that intertidal sediments had more abundant dissolved oxygen and stronger bioturbation than the distant sea sediments, and compared with Illumina sequencing, 454 pyrosequencing results were more prone to higher mismatch rates and the emergence of artificial amplification products (Gomez-Alvarez et al., 2009; Quince et al., 2009).
A variety of taxa were detected at all sampling stations; however, Proteobacteria were the most abundant and extensive in sediments. These findings were consistent with results from the Hangzhou Bay (Su et al., 2018), Jiaozhou Bay (Zhao et al., 2014) and the Changjiang (Yangtze) River Estuary (Feng et al., 2009). In the current study, the average relative abundances of γ-proteobacteria, δ-proteobacteria and α-proteobacteria were 24.6%, 22.9% and 4.7%, respectively. Bacterial community structure was similar to that reported from previous studies of coastal marine sediments (Guan et al., 2014; Lv et al., 2016). At Sta. S69 near the Huanghe River Estuary, γ-proteobacteria and α-proteobacteria accounted for high relative abundances of 30.2% and 8.5%, respectively, while δ-proteobacteria accounted for only 10.6%. Studies have found that the input of freshwater at the Changjiang River Estuary impacts the microbial community in the estuarine sediments (Guo et al., 2018). The relative abundance indicated that the bacterial community structure in S69 sediments with continuous freshwater and sediment input from the Huanghe River Estuary was significantly different from that in marine sediments. Higher levels of sediment in the southern region originated from the Huanghe River, and the impact of freshwater input on bacterial community structure provided a possible explanation for this difference. The current study also revealed that there were a large number of diverse microbial resources in the semi-open seas of the Bohai Sea.
The distribution pattern of bacteria was mainly driven by two factors, geographical distance and environmental variables. There was an obvious negative correlation between the distribution of bacterial communities and geographical distance (Martiny et al., 2011). Geographical barriers, such as ocean currents, water masses, and depth (pressure) (Brown et al., 2009; Zhang et al., 2014), constrain bacterial spread, and so the effects of constructing bacterial communities at small spatial scales such as the Bohai Sea were significant. Studies have found that Firmicutes occupy an important role in coastal sediments (Zinger et al., 2011). However, the average abundance of Firmicutes in sediments of the Bohai Sea was found to be 0.35%, far lower than the relative abundance of Bacteroidetes. Bacteroidetes accounted for 11.8% of the surface sediment in the Bohai Sea, and may play a role in the degradation of molecular organic matter in the environment (Sinkko et al., 2013). The discovery of Chloroflex (5.0%) at the 13 sampling stations means that sediments in the Bohai Sea were subjected to oil pollution, as it has been noted in previous studies that the bacteria within this phylum play a key role in petroleum degradation (Woese, 1987). Therefore, although the Bohai Sea is a geographically small area, it comprises various ecosystems, and the microbial community diversity of the area is a rich resource, which warrants further study. The microbial processes lay a solid foundation for revealing the geochemical cycles in which microbes participate.
In addition to environmental factors, the presence of pollutants (including oil pollution and heavy metal pollution) at various concentrations in marine sediments can also have significant effects on bacterial community structure (Wang and Tam, 2012). Therefore, the analysis of bacterial community structure is of great significance, because it can indirectly reflect multiple changes in the marine environment. Desulfuromonadales of δ-proteobacteria, detected in the current study, were often found in an environment under metal stress, which means that they have metal reducing ability (Cabrera et al., 2006; Vandieken et al., 2012). In the current study, Desulfuromonadales had an average abundance of 2.83% overall; however, the relative abundance of Desulfuromonadales at Stas S69, S104, C63, C83 and N79, which are more seriously polluted by heavy metals, was lower than others. Stations S101, C64 and C73 had relatively unpolluted sediments. Xanthomonadales was the dominant order, accounting for 12.28% of all orders. This order is generally detectable in marine sediments; however, these organisms use various other carbon substrates as carbon sources, and are classified as a general hydrocarbon degrading microorganisms (Gutierrez, 2017). Therefore, it is possible that the high abundance in the northern region may be due to a higher oil content in the sediment than in the central and southern regions. Desulfobacterales in the δ-Proteobacteria were the second most abundant order, accounting for an average relative abundance of 9.23%. Desulfobacterales may play a role in the sulfate reduction process, indicating that sulfate reduction processes are common in sediments of the Bohai Sea. These sediments are mainly affected by human activities. These results may reveal that the organic pollution caused by the industrial and agricultural development of the surrounding coastal cities in the Bohai Sea environment was significant and different, with contamination of regional sediments significantly affecting patterns of microbial community composition.
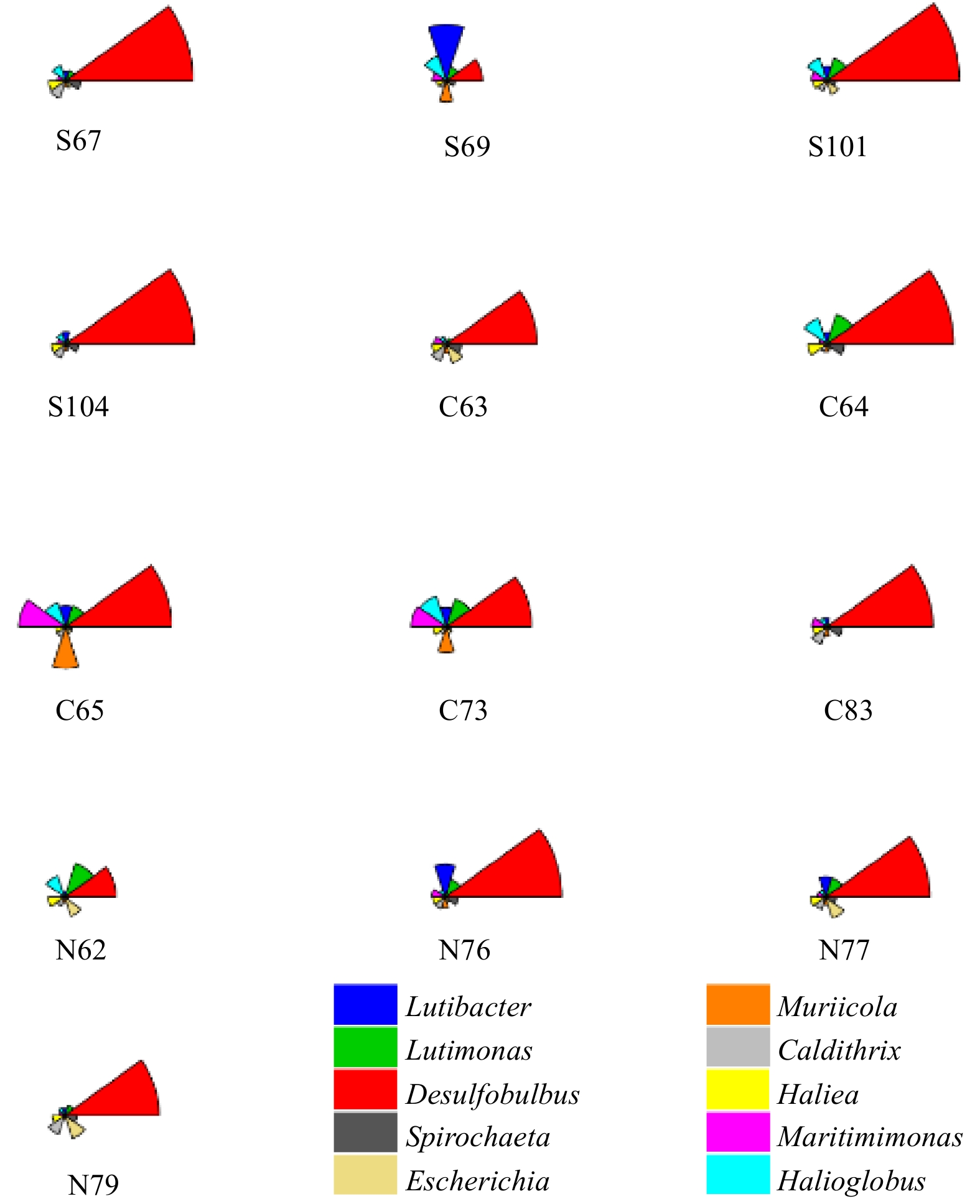
Desulfobulbus was the most abundant genus at most sampling stations, as shown in Fig. 8. All members of this genus can reduce sulfate and thiosulfate to hydrogen sulfide gas (Kuever et al., 2015). Among the dominant species, Lutimonas, Halioglobus and Caldithrix were significantly associated with Cr, Cu, Pb, Cd and Zn (P<0.05). Some Lutimonas have been found to contain catalase and oxidase, and have the ability to reduce nitrate to nitrite (Kim et al., 2014). In the ocean, the genus Halioglobus is found to reduce nitrate to nitrogen (Park et al., 2012). Caldithrix is a common genus in the ocean, which is associated with anaerobic respiration with molecular hydrogen or acetate as an electron donor and nitrate as an electron acceptor, especially in thermophilic, halophilic and sulfide-rich environments. It can be present in large amounts in petroleum-containing sediments and participates in the nitrogen cycle, forming ammonium salts (Alauzet and Jumas-Bilak, 2014; Miroshnichenko et al., 2003). In the current study, it was also found that Caldithrix was significantly correlated with sediment particle size (P<0.01) and TOC (P<0.05). At Stas S104 and S101, which were most polluted by Hg, the relative abundance of the Eudoraea genus was significantly higher than that of other stations. As one of the dominant genera, it is speculated that the genus may have a better Hg tolerance. Some studies found that the Eudoraea genus has no effect on nitrates, but can use various carbon compounds as the sole carbon source (Alain et al., 2008). The sediment of sampling Sta. 83 was found to be polluted by various heavy metals. The relative abundance of Desulfoconvexum was found to be four times higher at Sta. 83 than the average relative abundance of other stations, and was found to be abundant in the southern region where heavy metal pollution was relatively serious. It was significantly correlated with nitrate (P<0.05). Within this genus, some species were found to be able to utilize organic matter in a variety of environments and eventually completely oxidized to carbon dioxide (Könneke et al., 2013).

(1) The main groups of bacteria in the sediments of the Bohai Sea included Proteobacteria, Bacteroidetes, Planctomycetes, Acidobacteria and Chloroflexi. At the genus level, Desulfobulbus was most abundant, followed by Lutimonas, Halioglobus and Caldithrix, with abundance significantly related to the presence of heavy metals.
(2) Using real-time PCR and high-throughput sequencing of bacterial 16S rRNA genes in sediment samples from 13 sampling stations in the Bohai Sea, community structure diversity and abundance of bacteria in the sediment were found to increase from the northern region to the central region, and further increase in the southern region.
(3) In the Bohai Sea sediments were mainly polluted by Hg, with highest levels of Hg pollution in the southern region, particularly at Stas S101 and S104, where the Hg the Hg concentration was extremely high, an issue which deserves attention. In addition, sediments in the sea around Qinhuangdao were also affected by a variety of heavy metals.
(4) The main factors influencing bacterial community structure in the sediments of the Bohai Sea were TOC content in the sediment, followed by the depth and TP. The correlation between heavy metals and bacterial communities showed the consistency of the effects of Pb, Cd, Cr, Cu and Zn on the community, while the effect of Hg on the community structure was not significant. Arsenic content also showed a negative correlation with bacterial community structure in the majority of samples.
| [1] |
Alain K, Intertaglia L, Catala P, et al. 2008. Eudoraea adriatica gen. nov., sp. nov., a novel marine bacterium of the family Flavobacteriaceae. International Journal of Systematic and Evolutionary Microbiology, 58(10): 2275–2281. doi: 10.1099/ijs.0.65446-0
|
| [2] |
Alauzet C, Jumas-Bilak E. 2014. The phylum deferribacteres and the genus Caldithrix. In: Rosenberg E, DeLong E F, Lory S, et al., eds. The Prokaryotes. Berlin, Heidelberg: Springer, 595–611
|
| [3] |
Brown M V, Philip G K, Bunge J A, et al. 2009. Microbial community structure in the North Pacific Ocean. The ISME Journal, 3(12): 1374–1386. doi: 10.1038/ismej.2009.86
|
| [4] |
Cabrera G, Pérez R, Gómez J M, et al. 2006. Toxic effects of dissolved heavy metals on Desulfovibrio vulgaris and Desulfovibrio sp. strains. Journal of Hazardous Materials, 135(1–3): 40–46. doi: 10.1016/j.jhazmat.2005.11.058
|
| [5] |
Dai Tianjiao, Zhang Yan, Tang Yushi, et al. 2016. Identifying the key taxonomic categories that characterize microbial community diversity using full-scale classification: a case study of microbial communities in the sediments of Hangzhou Bay. FEMS Microbiology Ecology, 92(10): fiw150. doi: 10.1093/femsec/fiw150
|
| [6] |
Fan Jingfeng, Chen jiaying, Chen Liguang, et al. 2011. Research on denitrifying bacteria quantification and diversity in Liaohe Estuary sediments. Haiyang Xuebao (in Chinese), 33(3): 94–102
|
| [7] |
Feng Biwei, Li Xiaoran, Wang Jinhui, et al. 2009. Bacterial diversity of water and sediment in the Changjiang estuary and coastal area of the East China Sea. FEMS Microbiology Ecology, 70(2): 236–248. doi: 10.1111/j.1574-6941.2009.00772.x
|
| [8] |
General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of China. 2008. GB 17378.5-2007 The specification for marine monitoring—Part 5: sediment analysis (in Chinese). Beijing: Standards Press of China, 1–104
|
| [9] |
Gomez-Alvarez V, Teal T K, Schmidt T M. 2009. Systematic artifacts in metagenomes from complex microbial communities. The ISME Journal, 3(11): 1314–1317. doi: 10.1038/ismej.2009.72
|
| [10] |
Guan Xiangyu, Zhu Lingling, Li Youxun, et al. 2014. Composition and variation of sediment bacterial and nirS-harboring bacterial communities at representative sites of the Bohai Gulf coastal zone, China. World Journal of Microbiology and Biotechnology, 30(4): 1291–1300. doi: 10.1007/s11274-013-1553-4
|
| [11] |
Guo Xingpan, Lu Dapei, Niu Zuoshun, et al. 2018. Bacterial community structure in response to environmental impacts in the intertidal sediments along the Yangtze Estuary, China. Marine Pollution Bulletin, 126: 141–149. doi: 10.1016/j.marpolbul.2017.11.003
|
| [12] |
Gutierrez T. 2017 Aerobic Hydrocarbon-degrading Gammaproteobacteria: Xanthomonadales. In: McGenity T, ed. Taxonomy, Genomics and Ecophysiology of Hydrocarbon-Degrading Microbes. Cham: Springer, 1–15
|
| [13] |
Jing Xiaoyan, Gou Honglei, Gong Yanhai, et al. 2018. Raman-activated cell sorting and metagenomic sequencing revealing carbon-fixing bacteria in the ocean. Environmental Microbiology, 20(6): 2241–2255. doi: 10.1111/1462-2920.14268
|
| [14] |
Kim Y O, Park S, Nam B H, et al. 2014. Description of Lutimonas halocynthiae sp. nov., isolated from a golden sea squirt (Halocynthia aurantium), reclassification of Aestuariicola saemankumensis as Lutimonas saemankumensis comb. nov. and emended description of the genus Lutimonas. International Journal of Systematic and Evolutionary Microbiology, 64(6): 1984–1990
|
| [15] |
Könneke M, Kuever J, Galushko A, et al. 2013. Desulfoconvexum algidum gen. nov., sp. nov., a psychrophilic sulfate-reducing bacterium isolated from a permanently cold marine sediment. International Journal of Systematic and Evolutionary Microbiology, 63(3): 959–964
|
| [16] |
Köster M, Meyer-Reil L A. 2001. Characterization of carbon and microbial biomass pools in shallow water coastal sediments of the southern Baltic Sea (Nordrügensche Bodden). Marine Ecology Progress Series, 214: 25–41. doi: 10.3354/meps214025
|
| [17] |
Kuever J, Rainey F A, Widdel F. 2015. Desulfobulbus. In: Whitman W B, DeVos P, Chun J, et al., eds. Bergey’s Manual of Systematics of Archaea and Bacteria. Hoboken: John Wiley & Sons, Inc, 1–6
|
| [18] |
Li Hongjun, Ye Sheng, Ye Jinqing, et al. 2017. Baseline survey of sediments and marine organisms in Liaohe Estuary: heavy metals, polychlorinated biphenyls and organochlorine pesticides. Marine Pollution Bulletin, 114(1): 555–563. doi: 10.1016/j.marpolbul.2016.09.002
|
| [19] |
Liu Zongzhi, DeSantis T Z, Andersen G L, et al. 2008. Accurate taxonomy assignments from 16S rRNA sequences produced by highly parallel pyrosequencers. Nucleic Acids Research, 36(18): e120. doi: 10.1093/nar/gkn491
|
| [20] |
Liu Jiwen, Liu Xiaoshou, Wang Min, et al. 2015. Bacterial and archaeal communities in sediments of the north Chinese marginal seas. Microbial Ecology, 70(1): 105–117. doi: 10.1007/s00248-014-0553-8
|
| [21] |
Lofrano G, Libralato G, Minetto D, et al. 2017. In situ remediation of contaminated marinesediment: an overview. Environmental Science and Pollution Research, 24(6): 5189–5206. doi: 10.1007/s11356-016-8281-x
|
| [22] |
Lv Xiaofei, Ma Bin, Yu Junbao, et al. 2016. Bacterial community structure and function shift along a successional series of tidal flats in the Yellow River Delta. Scientific Reports, 6: 36550. doi: 10.1038/srep36550
|
| [23] |
Marshall I P G, Karst S M, Nielsen P H, et al. 2018. Metagenomes from deep Baltic Sea sediments reveal how past and present environmental conditions determine microbial community composition. Marine Genomics, 37: 58–68. doi: 10.1016/j.margen.2017.08.004
|
| [24] |
Martiny J B H, Eisen J A, Penn K, et al. 2011. Drivers of bacterial β-diversity depend on spatial scale. Proceedings of the National Academy of Sciences of the United States of America, 108(19): 7850–7854. doi: 10.1073/pnas.1016308108
|
| [25] |
Ministry of Environmental Protection of the People’s Republic of China. 2012. HJ 634-2012 Soil-determination of ammonium, nitrite and nitrate by extraction with potassium chloride solution-spectrophotometric methods (in Chinese). Beijing: China Environmental Press, 1–18
|
| [26] |
Miroshnichenko M L, Kostrikina N A, Chernyh N A, et al. 2003. Caldithrix abyssi gen. nov., sp. nov., a nitrate-reducing, thermophilic, anaerobic bacterium isolated from a Mid-Atlantic Ridge hydrothermal vent, represents a novel bacterial lineage. International Journal of Systematic and Evolutionary Microbiology, 53(1): 323–329. doi: 10.1099/ijs.0.02390-0
|
| [27] |
Muller G. 1969. Index of geoaccumulation in sediments of the Rhine River. Geojournal, 2(3): 108–118
|
| [28] |
Müller G, Förstner U. 1976. Schwermetalle in den Sedimenten der Elbe bei Stade: veränderungen seit 1973. Naturwissenschaften, 63(5): 242–243. doi: 10.1007/BF00610914
|
| [29] |
Park S, Yoshizawa S, Inomata K, et al. 2012. Halioglobus japonicus gen. nov., sp. nov. and Halioglobus pacificus sp. nov., members of the class Gammaproteobacteria isolated from seawater. International Journal of Systematic and Evolutionary Microbiology, 62(8): 1784–1789
|
| [30] |
Quince C, Lanzén A, Curtis T P, et al. 2009. Accurate determination of microbial diversity from 454 pyrosequencing data. Nature Methods, 6(9): 639–641. doi: 10.1038/nmeth.1361
|
| [31] |
Shade A. 2018. Understanding microbiome stability in a changing world. Msystems, 3(2): e00157-17. doi: 10.1128/mSystems.00157-17
|
| [32] |
Sinkko H, Lukkari K, Sihvonen L M, et al. 2013. Bacteria contribute to sediment nutrient release and reflect progressed eutrophication-driven hypoxia in an organic-rich continental sea. PLoS One, 8(6): e67061. doi: 10.1371/journal.pone.0067061
|
| [33] |
Su Zhiguo, Dai Tianjiao, Tang Yushi, et al. 2018. Sediment bacterial community structures and their predicted functions implied the impacts from natural processes and anthropogenic activities in coastal area. Marine Pollution Bulletin, 131: 481–495. doi: 10.1016/j.marpolbul.2018.04.052
|
| [34] |
Tian Qi, Wang Jia, Fan Xiaolei, et al. 2014. Methanogenic activity and methanogen diversity in marine gas field sediments. Environmental Science (in Chinese), 35(6): 2322–2327
|
| [35] |
Vandieken V, Pester M, Finke N, et al. 2012. Three manganese oxide-rich marine sediments harbor similar communities of acetate-oxidizing manganese-reducing bacteria. The ISME Journal, 6(11): 2078–2090. doi: 10.1038/ismej.2012.41
|
| [36] |
Wada M, Suzuki S, Nara T, et al. 2012. Microbial community respiration and structure of dead zone sediments of Omura Bay, Japan. Journal of Oceanography, 68(6): 857–867. doi: 10.1007/s10872-012-0136-6
|
| [37] |
Wang Haitao, Gilbert J A, Zhu Yongguan, et al. 2018. Salinity is a key factor driving the nitrogen cycling in the mangrove sediment. Science of the Total Environment, 631–632: 1342–1349. doi: 10.1016/j.scitotenv.2018.03.102
|
| [38] |
Wang Yafen, Tam N F Y. 2012. Natural attenuation of contaminated marine sediments from an old floating dock Part II: changes of sediment microbial community structure and its relationship with environmental variables. Science of the Total Environment, 423: 95–103. doi: 10.1016/j.scitotenv.2012.01.066
|
| [39] |
Woese C R. 1987. Bacterial evolution. Microbiological Reviews, 51(2): 221–271
|
| [40] |
Yang Yonghua, Yao Jian, Hua Xiaomei. 2000. Effect of pesticide pollution against functional microbial diversity in soil. Journal of Microbiology (in Chinese), 20(2): 23–25, 27
|
| [41] |
Zhang Huizhen, Chang Yongkai, Chen Quanrui, et al. 2018. Community structure analysis of Archaea and bacteria in sediments of Liaohe Estuary. Haiyang Xuebao (in Chinese), 40(6): 113–130
|
| [42] |
Zhang Yao, Zhao Zihao, Dai Minhan, et al. 2014. Drivers shaping the diversity and biogeography of total and active bacterial communities in the South China Sea. Molecular Ecology, 23(9): 2260–2274. doi: 10.1111/mec.12739
|
| [43] |
Zhao Yangguo, Feng Gong, Bai Jie, et al. 2014. Effect of copper exposure on bacterial community structure and function in the sediments of Jiaozhou Bay, China. World Journal of Microbiology and Biotechnology, 30(7): 2033–2043. doi: 10.1007/s11274-014-1628-x
|
| [44] |
Zheng Binghui, Wang Liping, Liu Lusan. 2014. Bacterial community structure and its regulating factors in the intertidal sediment along the Liaodong Bay of Bohai Sea, China. Microbiological Research, 169(7–8): 585–592. doi: 10.1016/j.micres.2013.09.019
|
| [45] |
Zhou Kan, Fan Jie, Liu Hanchu. 2017. Spatiotemporal patterns and driving forces of water pollutant discharge in the Bohai Rim Region. Progress in Geography (in Chinese), 36(2): 171–181. doi: 10.18306/dlkxjz.2017.02.004
|
| [46] |
Zinger L, Amaral-Zettler L A, Fuhrman J A, et al. 2011. Global patterns of bacterial beta-diversity in seafloor and seawater ecosystems. PLoS One, 6(9): e24570. doi: 10.1371/journal.pone.0024570
|
| 1. | Yuting Yang, Qiwen Liang, Yisheng Peng, et al. Differential mediation of biogeochemical processes through bioturbation by fiddler and sesarmid mangrove crabs. Marine Pollution Bulletin, 2025, 211: 117431. doi:10.1016/j.marpolbul.2024.117431 | |
| 2. | Ileana Sarahi Ramos-Mendoza, Salvador Embarcadero-Jiménez, Andrea Fernanda Barrios-Navarro, et al. Prokaryotic community structure and predicted metabolism associated with hydrocarbon degradation in marine sediments from the Northwest Coast of Baja California, Mexico. Journal of Soils and Sediments, 2024, 24(8): 3148. doi:10.1007/s11368-024-03822-w | |
| 3. | Wenchao Deng, Shunyang Chen, Shiquan Chen, et al. Impacts of eutrophication on microbial community structure in sediment, seawater, and phyllosphere of seagrass ecosystems. Frontiers in Microbiology, 2024, 15 doi:10.3389/fmicb.2024.1449545 | |
| 4. | Lilian Wen, Jiajia Dai, Jinming Song, et al. Antibiotic resistance genes (ARGs) and their eco-environmental response in the Bohai Sea sediments. Marine Pollution Bulletin, 2024, 208: 116979. doi:10.1016/j.marpolbul.2024.116979 | |
| 5. | Ming Hongxia, Zan Shuaijun, Liu Jiwen, et al. Promoting the denitrification process by heavy metals in Liaohe Estuary sediment. Marine Pollution Bulletin, 2024, 203: 116408. doi:10.1016/j.marpolbul.2024.116408 | |
| 6. | Huili Zeng, Jingyi Huang, Minghuo Wu, et al. Pyrene biodegradation by bacterial consortia enriched from coastal and marine sediments: Insights into bacterial community structures and functions. Journal of Environmental Chemical Engineering, 2024, 12(5): 113922. doi:10.1016/j.jece.2024.113922 | |
| 7. | Zhansheng Guo, Wang Lu, Song Minpeng, et al. Seasonal dynamics response mechanism of benthic microbial community to artificial reef habitats. Environmental Research, 2024, 243: 117867. doi:10.1016/j.envres.2023.117867 | |
| 8. | Changfeng Qu, Xixi Wang, Liping Zhang, et al. Prokaryotic diversity and community composition in the surface sediments of the Changjiang River Estuary in summer. Acta Oceanologica Sinica, 2023, 42(9): 113. doi:10.1007/s13131-023-2153-2 | |
| 9. | Zimin Wei, Yue Zhao, Li Zhao, et al. The contribution of microbial shikimic acid to humus formation during organic wastes composting: a review. World Journal of Microbiology and Biotechnology, 2023, 39(9) doi:10.1007/s11274-023-03674-5 | |
| 10. | Xiangfeng Zeng, Yongbin Li, Shaofeng Wang, et al. Diversity, functional potential, and assembly of bacterial communities in metal(loid)-contaminated sediments from a coastal industrial basin. Science of The Total Environment, 2023, 886: 163831. doi:10.1016/j.scitotenv.2023.163831 | |
| 11. | Yan Zhang, Jingjing Liu, Lei Kang, et al. Temporal variation of mineralization rates and its influence on carbon storage over the last 50 years in Bohai Bay, China. Marine Pollution Bulletin, 2023, 188: 114624. doi:10.1016/j.marpolbul.2023.114624 | |
| 12. | Tingxuan An, Xiaolan Lu, Yangchun Han, et al. Exploring the bacterial diversity and composition with special emphasis on pathogens in ship ballast water and sediments using full-length 16S rRNA gene sequencing. Marine Pollution Bulletin, 2023, 194: 115336. doi:10.1016/j.marpolbul.2023.115336 | |
| 13. | Jun-Feng Wang, Huan-Zhan Zhou, Bo Jiang, et al. Controlling heavy metals pollution is vital for the restoration of carbon and nitrogen transformation function of mangrove ecosystems in the Greater Bay Area, China. Journal of Water Process Engineering, 2023, 53: 103745. doi:10.1016/j.jwpe.2023.103745 | |
| 14. | Yanan Cheng, Qiongyuan Su, Shu Yang, et al. Sedimentary bacterial communities in subtropical Beibu Gulf: Assembly process and functional profile. Ecological Indicators, 2023, 154: 110814. doi:10.1016/j.ecolind.2023.110814 | |
| 15. | Caixia Wang, Jiujun Ju, Haikun Zhang, et al. Disclosing the ecological implications of heavy metal disturbance on the microbial N-transformation process in the ocean tidal flushing urban estuary. Ecological Indicators, 2022, 144: 109504. doi:10.1016/j.ecolind.2022.109504 | |
| 16. | Xianzhe Gong, Zhiyi Chen, Ye Deng, et al. Contrasting archaeal and bacterial community assembly processes and the importance of rare taxa along a depth gradient in shallow coastal sediments. Science of The Total Environment, 2022, 852: 158411. doi:10.1016/j.scitotenv.2022.158411 | |
| 17. | Jinmei Liu, Xiaolei Wang, Jiao Liu, et al. Comparison of assembly process and co-occurrence pattern between planktonic and benthic microbial communities in the Bohai Sea. Frontiers in Microbiology, 2022, 13 doi:10.3389/fmicb.2022.1003623 | |
| 18. | Ming Hongxia, Fan Jingfeng, Liu Jiwen, et al. Full-length 16S rRNA gene sequencing reveals spatiotemporal dynamics of bacterial community in a heavily polluted estuary, China. Environmental Pollution, 2021, 275: 116567. doi:10.1016/j.envpol.2021.116567 | |
| 19. | Quanrui Chen, Jingfeng Fan, Hongxia Ming, et al. Effects of environmental factors on denitrifying bacteria and functional genes in sediments of Bohai Sea, China. Marine Pollution Bulletin, 2020, 160: 111621. doi:10.1016/j.marpolbul.2020.111621 | |
| 20. | Qi Li, Min Chen, Renming Jia, et al. Transit time of river water in the Bering and Chukchi Seas estimated from δ18O and radium isotopes. Progress in Oceanography, 2017, 159: 115. doi:10.1016/j.pocean.2017.08.004 | |
| 21. | Wu Men, Yuwu Jiang, Guangshan Liu, et al. Study of water mixing in the coastal waters of the western Taiwan Strait based on radium isotopes. Journal of Environmental Radioactivity, 2016, 152: 16. doi:10.1016/j.jenvrad.2015.11.003 | |
| 22. | Wu Men, Fenfen Wang, Yusheng Zhang, et al. Determining coastal mixing rates of western Taiwan Strait using 224Ra. Journal of Radioanalytical and Nuclear Chemistry, 2013, 295(1): 89. doi:10.1007/s10967-012-1853-4 |
Figures(8) / Tables(2)

Supported by:
Beijing Renhe Information Technology Co. Ltd

Quanrui Chen, Jingfeng Fan, Jie Su, Hongxia Ming, Zhihao Sun, Mengfei Li, Xiaohui Zhao, Yantao Wang, Yingxue Zhang, Huizhen Zhang, Yuan Jin, Xiaowan Ma, Bin Wang. Spatial distribution characteristics of bacterial community structure and gene abundance in sediments of the Bohai Sea[J]. Acta Oceanologica Sinica, 2020, 39(2): 69-78. doi: 10.1007/s13131-020-1554-8

| Station | NO3–/μg·g–1 | NO2–/μg·g–1 | NH4+/μg·g–1 | TOC/% | TP/μg·g–1 | TN/μg·g–1 | depth/m | Grid size (Mz, Φ) |
| S67 | 421 | 29.9 | 153 | 0.66 | 143 | 130 | 23 | 6.63 |
| S69 | 513 | 27.6 | 173 | 0.43 | 386 | 122 | 10 | 5.76 |
| S101 | 482 | 59 | 140 | 0.36 | 174 | 93.6 | 27 | 5.64 |
| S104 | 256 | 29.9 | 121 | 0.53 | 348 | 135 | 17.5 | 6.11 |
| C63 | 217 | 68.9 | 96.2 | 0.49 | 255 | 97.3 | 28 | 6.22 |
| C64 | 149 | 85.3 | 101 | 0.24 | 181 | 91.6 | 19.8 | 4.99 |
| C65 | 228 | 70.7 | 104 | 0.39 | 208 | 93.6 | 19 | 4.64 |
| C73 | 183 | 103 | 68.3 | 0.62 | 221 | 145 | 34 | 6.21 |
| C83 | 330 | 83.4 | 93.3 | 0.92 | 269 | 157 | 23 | 7.37 |
| N62 | 185 | 75 | 84.9 | 0.36 | 212 | 95.8 | 26 | 5.43 |
| N76 | 350 | 75.4 | 112 | 0.84 | 290 | 114 | 13 | 6.3 |
| N77 | 256 | 67.4 | 72 | 0.53 | 219 | 100 | 12.7 | 5.81 |
| N79 | 138 | 58.5 | 197 | 0.75 | 245 | 111 | 12.5 | 6.52 |
DownLoad:
CSV
| Station | Reads | OTU | Ace | Chao1 | Shannon | Simpson | Coverage/% |
| S67 | 28 770 | 2 182 | 2 547 | 2 520 | 6.13 | 0.008 2 | 98.2 |
| S69 | 22 563 | 1 585 | 2 029 | 2 079 | 5.83 | 0.009 9 | 97.9 |
| S101 | 32 385 | 1 963 | 2 299 | 2 328 | 5.94 | 0.008 7 | 98.6 |
| S104 | 22 514 | 2 017 | 2 395 | 2 397 | 6.1 | 0.008 6 | 97.7 |
| C63 | 26 467 | 2 011 | 2 371 | 2 347 | 6.07 | 0.008 8 | 98.2 |
| C64 | 24 761 | 1 812 | 2 245 | 2 208 | 5.59 | 0.016 1 | 97.9 |
| C65 | 25 325 | 1 825 | 2 282 | 2 248 | 5.78 | 0.009 8 | 98.0 |
| C73 | 28 309 | 1 838 | 2 345 | 2 370 | 5.62 | 0.014 7 | 98.1 |
| C83 | 24 577 | 2 117 | 2 473 | 2 505 | 6.24 | 0.007 2 | 98.0 |
| N62 | 24 563 | 1 727 | 2 135 | 2 113 | 5.78 | 0.010 8 | 98.1 |
| N76 | 26 143 | 2 065 | 2 457 | 2 428 | 6.07 | 0.009 2 | 98.0 |
| N77 | 24 527 | 1 892 | 2 325 | 2 279 | 5.92 | 0.011 3 | 97.9 |
| N79 | 26 312 | 1 873 | 2 257 | 2 239 | 5.93 | 0.009 3 | 98.2 |
DownLoad:
CSV
| Station | NO3–/μg·g–1 | NO2–/μg·g–1 | NH4+/μg·g–1 | TOC/% | TP/μg·g–1 | TN/μg·g–1 | depth/m | Grid size (Mz, Φ) |
| S67 | 421 | 29.9 | 153 | 0.66 | 143 | 130 | 23 | 6.63 |
| S69 | 513 | 27.6 | 173 | 0.43 | 386 | 122 | 10 | 5.76 |
| S101 | 482 | 59 | 140 | 0.36 | 174 | 93.6 | 27 | 5.64 |
| S104 | 256 | 29.9 | 121 | 0.53 | 348 | 135 | 17.5 | 6.11 |
| C63 | 217 | 68.9 | 96.2 | 0.49 | 255 | 97.3 | 28 | 6.22 |
| C64 | 149 | 85.3 | 101 | 0.24 | 181 | 91.6 | 19.8 | 4.99 |
| C65 | 228 | 70.7 | 104 | 0.39 | 208 | 93.6 | 19 | 4.64 |
| C73 | 183 | 103 | 68.3 | 0.62 | 221 | 145 | 34 | 6.21 |
| C83 | 330 | 83.4 | 93.3 | 0.92 | 269 | 157 | 23 | 7.37 |
| N62 | 185 | 75 | 84.9 | 0.36 | 212 | 95.8 | 26 | 5.43 |
| N76 | 350 | 75.4 | 112 | 0.84 | 290 | 114 | 13 | 6.3 |
| N77 | 256 | 67.4 | 72 | 0.53 | 219 | 100 | 12.7 | 5.81 |
| N79 | 138 | 58.5 | 197 | 0.75 | 245 | 111 | 12.5 | 6.52 |
| Station | Reads | OTU | Ace | Chao1 | Shannon | Simpson | Coverage/% |
| S67 | 28 770 | 2 182 | 2 547 | 2 520 | 6.13 | 0.008 2 | 98.2 |
| S69 | 22 563 | 1 585 | 2 029 | 2 079 | 5.83 | 0.009 9 | 97.9 |
| S101 | 32 385 | 1 963 | 2 299 | 2 328 | 5.94 | 0.008 7 | 98.6 |
| S104 | 22 514 | 2 017 | 2 395 | 2 397 | 6.1 | 0.008 6 | 97.7 |
| C63 | 26 467 | 2 011 | 2 371 | 2 347 | 6.07 | 0.008 8 | 98.2 |
| C64 | 24 761 | 1 812 | 2 245 | 2 208 | 5.59 | 0.016 1 | 97.9 |
| C65 | 25 325 | 1 825 | 2 282 | 2 248 | 5.78 | 0.009 8 | 98.0 |
| C73 | 28 309 | 1 838 | 2 345 | 2 370 | 5.62 | 0.014 7 | 98.1 |
| C83 | 24 577 | 2 117 | 2 473 | 2 505 | 6.24 | 0.007 2 | 98.0 |
| N62 | 24 563 | 1 727 | 2 135 | 2 113 | 5.78 | 0.010 8 | 98.1 |
| N76 | 26 143 | 2 065 | 2 457 | 2 428 | 6.07 | 0.009 2 | 98.0 |
| N77 | 24 527 | 1 892 | 2 325 | 2 279 | 5.92 | 0.011 3 | 97.9 |
| N79 | 26 312 | 1 873 | 2 257 | 2 239 | 5.93 | 0.009 3 | 98.2 |
 DownLoad:
DownLoad:



 DownLoad:
DownLoad: