Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture/Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
2.
Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China
Funds:
The National Key R&D Program of China under contract No. 2019YFD0900400; the China Agriculture Research System under contract No. CARS-48; the National Natural Science Foundation of China under contract No. 31702319; the Program of Shandong Leading Talent under contract No. LNJY2015002; the Central Public-interest Scientific Institution Basal Research Fund, CAFS under contract No. 2019ZD0603.
The ridgetail white prawn Exopalaemon carinicauda is a euryhaline shrimp species in the estuarine and coastal areas of China. In this study, survival rates, transcription levels of two prophenoloxidase system-related genes (EcLGBP and EcproPO) and PO activity were determined quantitatively in juvenile and adult E. carinicauda under different salinity levels. The results showed that E. carinicauda juveniles could survive in a wider range of salinity conditions than adults. For juvenile E. carinicauda, the expression levels of EcLGBP and EcProPO were up-regulated in low salinities and showed no significant difference at 20–40, while PO activities in low salinities were higher compared to those in high salinities. For adult E. carinicauda, the expression profiles of EcLGBP and EcproPO had a different trend of up-regulation in salinity stress treatments and no obvious difference was observed in the gene expression levels and PO activity between 30 and 40. The salinity tolerance range of immunity for juvenile and adult E. carinicauda is 20–40 and 30–40, respectively.
In recent decades, mass mortalities and great economic losses of shrimp have been documented, with numerous studies demonstrating that environmental factors, including temperature, salinity, dissolved oxygen and pH, can affect the immune system of shrimp (Pascual et al., 2003; Cheng and Chen, 2000; Chen et al., 2007). In particular, salinity changes have been known to suppress the physiological and immune system of shrimp, leading to an increased susceptibility to pathogens (de la Vega et al., 2007; Kumlu et al., 2000; Pan et al., 2007; Joseph and Philip, 2007; Cheng and Chen, 2000). Therefore, the research on the mechanism of immune regulation in shrimp under salinity changes is an urgent issue in shrimp aquaculture.
The shrimp defense system is categorized as innate immunity that includes circulating haemocytes and various active substances released into haemolymph such as the prophenoloxidase (proPO) system, antibacterial peptides, and lectins (Söderhäll and Cerenius 1992). Activation of the proPO system occurs due to the specific recognition of microorganisms (PRPs), triggering a serine proteinase cascade, eventually leading to the cleavage of the inactive proPO to the active phenoloxidase (PO) that functions to produce the melanin and toxic reactive intermediates against invading pathogens (Ishwarya et al., 2016). As important PRRs, lipopolysaccharide and β-1, 3-glucan binding protein (LGBP) has been documented to activate the prophenoloxidase system of shrimp (Lee et al., 2000; Cheng et al., 2005; Liu et al., 2009). The proPO system can either be specifically activated by microbial cell wall components including β-1, 3-glucan, lipopolysaccharide (LPS) and peptidoglycan (Amparyup et al., 2013; Johansson and Soderhall, 1989; Cerenius and Söderhäll, 2004; Cerenius et al., 2008; Li and Xiang, 2013; Yang et al., 2014) and can also be regulated by environmental or experimental factors including calcium, pH and temperature (Ai et al., 2008). Immune responses to salinity fluctuations can be well reflected in the proPO system. Phenoloxidase activity has been proved as an immune parameter under salinity stress in shrimp (Joseph and Philip, 2007; Cheng and Chen, 2000; Wang and Chen, 2006). Hu (2008) reported the effect of salinity changes on the proPO system of haemolymph in Litopenaeus vannamei. Recently, with the development of high-throughput next-generation sequencing techniques, Zhao et al. (2015) indicated that low salinity changes induced the proPO system in L. vannamei haemolymph by investigating the digital gene expression (DGE).
In general, salinity tolerance and immunity in shrimp are correlated with the developmental stages. Cieluch et al. (2005) reported that the adult pattern of osmoregulation in Crangon crangon is accomplished after metamorphosis from a moderately hyperosmoconforming decapodid to an effectively hyper-/hyporegulating juvenile stage and the salinity tolerance and osmoregulatory capacity are closely correlated with the development of ion-transporting cells and the expression of iontransporting enzymes. Jiravanichpaisal et al. (2007) found no functional proPO system was present during larval stages in Penaeus monodon and suggested that the proPO system was not transferred from the mother to the offspring. The shrimp develop the complete and effective hyper-/hyporegulating immune system gradually after the post-larval stage, and there was little comparison in osmoregulatory capacity and immunity between the juvenile and adult.
Due to good reproductive performance, fast growth and adaptability to a range of environmental conditions, farming of the ridgetail white prawn, Exopalaemon carinicauda, has been booming along the coasts of the Yellow Sea and Bohai Sea, China (Xu et al., 2010; Li et al., 2012, 2015; Zhang et al., 2014). Some studies have reported that E. carinicauda can tolerate a low salinity level of 4 to a high salinity level of 35, and its optimal range of salinity for growth and first sexual maturity is 10–15 (Wang et al., 2005, 2010). However, shrimp in different life stages are usually tolerant to different salinity ranges. To examine the salinity ranges that juvenile and adult E. carinicauda are tolerant to, survival of E. carinicauda under different salinity stress was recorded in this study. In addition, the majority of previous studies focused on the salinity stress of shrimp were usually conducted at moderate salinity levels, whereas extreme salinity conditions are more typical for transporting and culturing shrimp (Dong, 1989). Currently, little information is available on the influence of salinity changes on the immune regulation of E. carinicauda. To study the immune responses of extreme and moderate salinity stressed E. carinicauda, the transcript profiles of EcLGBP and EcproPO in the proPO system associated with PO activity were determined in this study. The study aimed to obtain the salinity tolerant ranges of E. carinicauda of different sizes and gain insight into the proPO system in E. carinicauda under salinity stress. This will provide not only a theoretical basis for understanding the proPO system regulation mechanism in the shrimp under salinity stress, but also a technical reference for evaluating immune responses in shrimp aquaculture.
2.
Materials and methods
2.1
Experimental design of salinity stress levels
A preliminary experiment was designed to confirm the tolerance limit of salinity by setting three low salinity groups (0, 1 and 3) and three high salinity groups (47, 49 and 50). Salinity was adjusted by adding fresh water or marine salts. Ultimately, the cumulative mortality rates under the lowest salinity 0 and the highest salinity 51 were almost 100% at 96 h. Then, ten salinity groups were designed for the survival experiment of juveniles and adults as follows: 1, 3, 6, 10, 20, 30 (control group), 40, 44, 47 and 49. For each group, there were three tanks used for three replicates, and each tank contained ten shrimp. The cumulative survival rates of the two shrimp sizes (juveniles and adults) were evaluated at different salinity levels. The cumulative survival rate was calculated at 24, 48, 72 and 96 h by the following formula: R = [S1+S2+…+St]/Nt × 100%, where R (%) indicates the survival rate, S indicates the number of surviving shrimp, t indicates the sampling time, and Nt indicates the total number of shrimp in every tank.
Six salinity groups were designed for the sampling experiments of juveniles and adults as follows: 1, 10, 20, 30 (control group), 40 and 49. For each group, there were three tanks used for three replicates, and each tank contained 50 shrimp. Because juveniles were too small to collect haemolymph, the whole individual was sampled. Six juvenile individuals and haemolymph from six adults were randomly sampled at 0, 24, 48, 72 and 96 h for each salinity group. For adults of each salinity group, 500 μL haemolymph was collected from the shrimp hearts with sterile syringes containing 500 μL anticoagulant solution (1.588 g sodium citrate, 3.92 g sodium chloride, 4.56 g glucose, 0.66 g EDTA-2Na, 200 mL ddH2O) (Söderhäll and Smith, 1983). Haemolymph was centrifuged at 3 000 r/min at 4°C for 10 min. The supernatant fluid was stored at –20°C for the measurement of PO activity, while the hemocytes were dissolved with Trizol Reagent (Invitrogen, USA) and stored at –80°C for RNA extraction. For juveniles, individuals were ground into powder in liquid nitrogen. The powder was divided into two parts: 0.05 g was dissolved in 450 μL PBS for the measurement of PO activity, and 0.05 g was dissolved in 500 μL Trizol Reagent for RNA extraction.
2.2
PO activity of juvenile and adult E. carinicauda under different salinity levels
Phenoloxidase activity was measured spectrophotometrically by recording the formation of dopachrome produced from L-dihydroxyphenylalanine (L-DOPA) based on the modified method by Ashida (1971) and Lei et al. (2001). The optical density of the shrimp’s PO activity at 490 nm was measured using an IMARK (model U-2000, Tokyo, Japan). Phenoloxidase activity was expressed as 200 μL 0.1 mol/L buffer solution of potassium phosphate (pH=6.4), followed by 10 μL 0.01 mol/L L-DOPA dopachrome formation per 10 μL of supernatant fluid. All samples for PO activity measure were replicated in triplicate. The activity was expressed as increase in absorbance per minute per 100 μL haemolymph.
2.3
Expression of candidate genes under different salinity levels
Quantitative real-time PCR was performed on an ABI PRISM 7500 Sequence Detection System (Applied Biosystems, USA) to investigate the expression patterns of candidate genes in juvenile and adult E. carinicauda at different salinity stress levels. The primers of the target genes are shown in Table 1. The 18S rRNA (GenBank accession number: GQ369794) of E. carinicauda was used as an internal control to verify the quantitative real-time PCR reaction for its stable expression in different tissues and times (Table 1). The qPCR amplifications were carried out in triplicate in a total volume of 20 μL containing 10 μL SYBR® Premix Ex TaqTM II (2×) (TaKaRa), 2 μL of the cDNA template, 0.8 μL each of F and R primer (10 mmol/L), 0.4 μL ROX Reference Dye II (50×) and 6.0 μL DEPC-treated water. The PCR program was 95°C for 30 s, then 40 cycles of 95°C for 5 s and 60°C for 34 s, followed by 1 cycle of 95°C for 15 s, 60°C for 1 min and 95°C for 15 s. The target expression levels at different salinity stress levels were calculated with 2–△△CT methods (Livak and Schmittgen, 2001; Duan et al., 2013). All of the data were presented as mean±SE and subjected to one-way analysis of variance followed by Duncan’s multiple range tests. Differences were considered significant when P<0.05.
3.1
Cumulative survival rates of juvenile and adult E. carinicauda under different salinity stress levels
Cumulative survival rates (%) are shown in Table 2 for juvenile E. carinicauda under different salinity stress levels. Almost all shrimp survived under 3–44 salinity stress levels. Eight or nine of ten shrimp reared at salinity 47 survived in the challenge experiment after 24 h. Only two or three of ten shrimp reared at salinity 49 survived after 48 h; the cumulative survival rates in the salinity 1 treatment group were significantly higher than the salinity 49 treatment group throughout the experimental period (P<0.05).
Table
2.
Cumulative survival rates of juvenile E. carinicauda after exposure to different salinity stress levels
Salinity
Time/h
24
48
72
96
1
86.67±5.77b
76.67±5.77b
73.33±5.77b
73.33±5.77b
3
100±0.00c
100±0.00c
100±0.00d
100±0.00d
6
100±0.00c
100±0.00c
100±0.00d
100±0.00d
10
100±0.00c
100±0.00c
100±0.00d
100±0.00d
20
100±0.00c
100±0.00c
100±0.00d
100±0.00d
30
100±0.00c
100±0.00c
100±0.00d
100±0.00d
40
100±0.00c
100±0.00c
100±0.00d
100±0.00d
44
96.67±5.77c
96.67±5.77c
96.67±5.77d
96.67±5.77d
47
83.33±5.77b
83.33±5.77b
83.33±5.77c
83.33±5.77c
49
46.67±5.77a
33.33±5.77a
23.33±5.77a
23.33±5.77a
Note: Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
Cumulative survival rates (%) of adult E. carinicauda exposed to different salinity conditions are shown in Table 3. The shrimp in the salinity range of 6–44 showed almost 100% survival rates. Seven or eight of ten shrimp reared at salinity 3 or 47 survived in the challenged experiment after 48 h and no significant difference was observed between the two treatment groups (P>0.05). The cumulative survival rate of adult shrimp reared at salinity 49 at 24 h was significantly higher than that at salinity 1 (P<0.05). Only three or four of ten shrimp reared at salinity 1 or 49 survived in the challenge experiment after 72 h, and no significant difference was observed between the two treatment groups after 48 h (P>0.05). In addition, for both juvenile and adult E. carinicauda, 100% survival rates of shrimp reared at salinity 10–40 were observed throughout the experimental period.
Table
3.
Cumulative survival rates of adult E. carinicauda after exposure to different salinity stress levels
Salinity
Time/h
24
48
72
96
1
53.33±5.77a
40±10.00a
33.33±5.77a
33.33±5.77a
3
86.67±5.77c
76.67±5.77b
73.33±5.77b
73.33±5.77b
6
96.67±5.77d
96.67±5.77c
96.67±5.77c
96.67±5.77c
10
100±0.00d
100±0.00c
100±0.00c
100±0.00c
20
100±0.00d
100±0.00c
100±0.00c
100±0.00c
30
100±0.00d
100±0.00c
100±0.00c
100±0.00c
40
100±0.00d
100±0.00c
100±0.00c
100±0.00c
44
96.67±5.77d
96.67±5.77c
96.67±5.77c
96.67±5.77c
47
83.33±5.77c
80.00±10.00b
76.67±5.77b
76.67±5.77b
49
63.33±5.77b
46.67±5.77a
40±0.00a
40±0.00a
Note: Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
3.2
Expression of EcLGBP gene in juvenile and adult E. carinicauda under low and high salinity stress
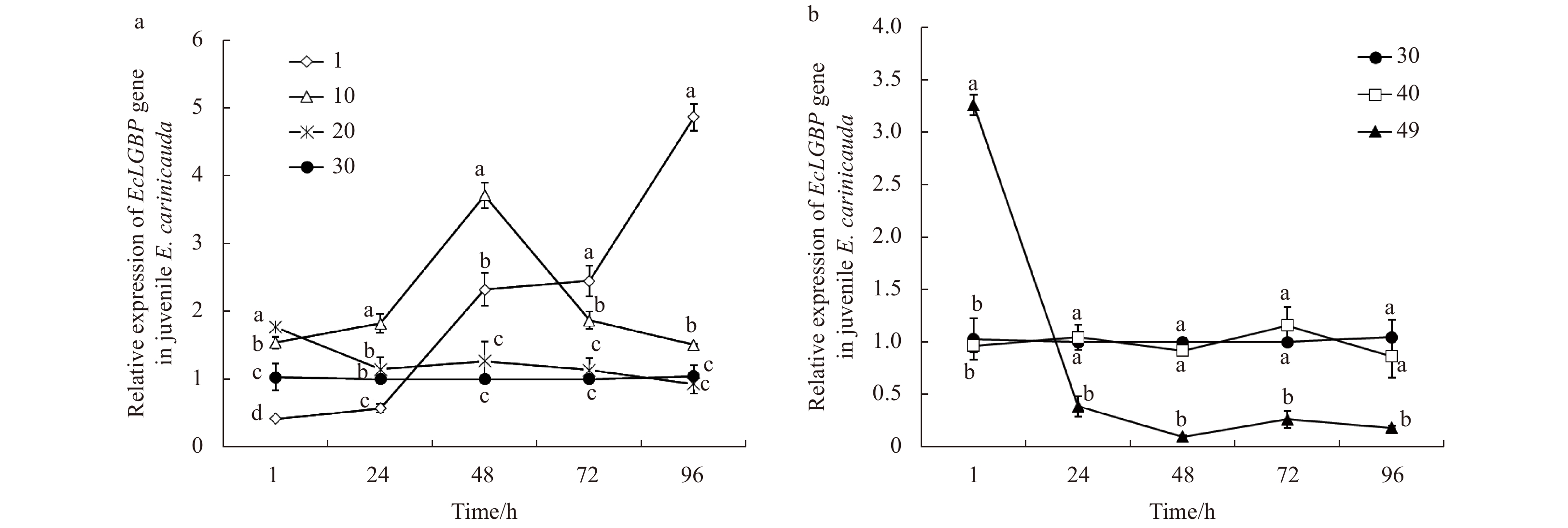
The expression profiles of the EcLGBP gene in juvenile E. carinicauda under low and high salinity stress are shown in Fig. 1. The transcriptional levels of EcLGBP in the salinity 1 group were down-regulated at first, and then the levels increased. The EcLGBP mRNA expression was up-regulated during the entire experimental period and reached a peak at 48 h in the salinity 10 group. No large change was observed at 24–96 h in the salinity 20 group (Fig. 1a). Under high salinity stress, the expression of EcLGBP was down-regulated in the salinity 49 group throughout the experimental period and reached the lowest level at 48 h. No significant changes in the expression of EcLGBP were observed in salinity 40 (P<0.05) (Fig. 1b).
Figure
1.
The expression profiles of the EcLGBP gene in juvenile E. carinicauda after low salinity (a) and high salinity (b) stress. Vertical bars represent the mean±SE (n=3). Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
The expression profiles of the EcLGBP gene in adult E. carinicauda under low and high salinity stress are shown in Fig. 2. Under low salinity stress levels, the expression levels of EcLGBP were up-regulated during the whole experimental period except 48 h in the salinity 1 group and reached a peak at 24 h and 72 h in both the salinity 10 and 20 groups (Fig. 2a). Under high salinity stress, the expression level of EcLGBP was up-regulated at 24 h and recovered to the control level after 48 h in the salinity 40 treatment group. In the salinity 49 group, the expression levels of EcLGBP fluctuated throughout the experimental period; the expression levels were up-regulated at 24 h, down-regulated at 48–72 h and then gradually up-regulated (Fig. 2b).
Figure
2.
The expression profiles of the EcLGBP gene in adult E. carinicauda after low salinity (a) and high salinity (b) stress. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
3.3
Expression of the EcproPO gene in juvenile and adult E. carinicauda under low and high salinity stress
The expression profiles of the EcproPO gene in juvenile E. carinicauda under low and high salinity stress are shown in Fig. 3. Under low salinity stress, the expression of EcproPO was up-regulated compared with the control group in the salinity 1 group throughout the experimental period except at 1 h, and the highest level was detected at 96 h. In the salinity 10 group, EcproPO mRNA expression was first up-regulated and then was down-regulated. In the salinity 20 group, the expression of EcproPO had no great change (Fig. 3a). Under high salinity stress, the expression levels of EcproPO were down-regulated at 24–48 h and then increased. In the salinity 49 treatment group, down-regulated expression of EcproPO was detected throughout the experimental period compared with the salinity 30 group (Fig. 3b).
Figure
3.
The expression profiles of the EcproPO gene in juvenile E. carinicauda after exposure to low salinity (a) and high salinity (b) stress. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
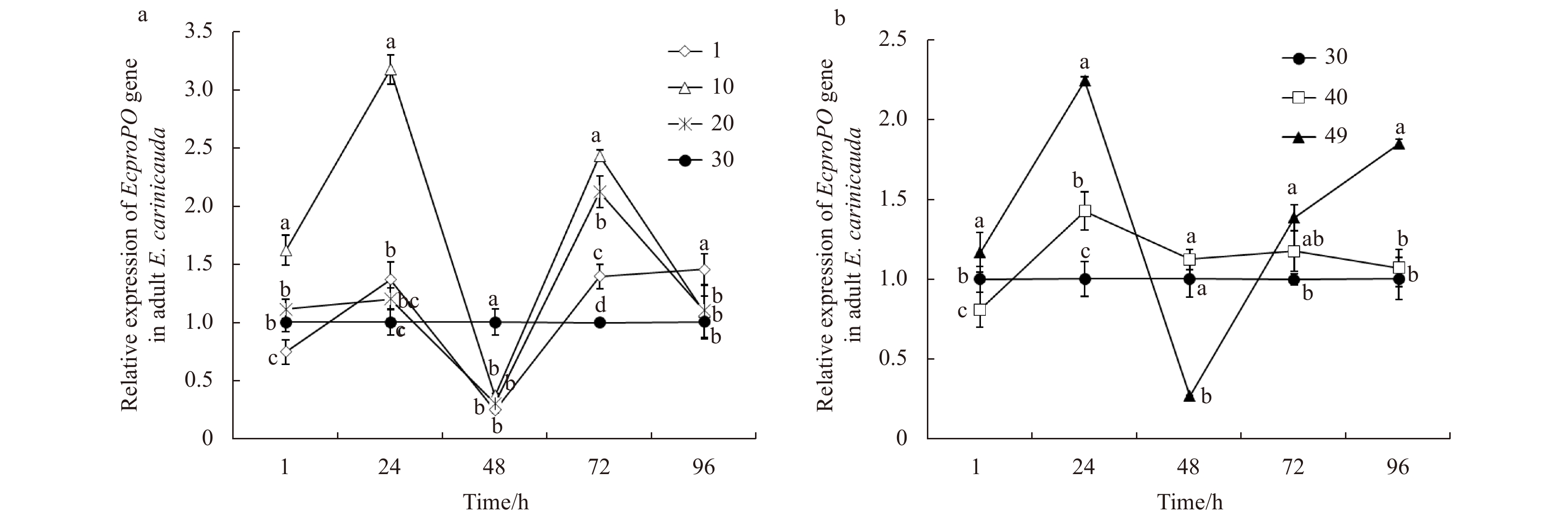
The expression profiles of the EcproPO gene in adult E. carinicauda under low and high salinity stress are shown in Fig. 4. Under low salinity stress, the expression profiles of EcproPO showed up-regulated and down-regulated changes with two-peaks at 24 h and 48 h, respectively (Fig. 4a). Under high salinity stress, mRNA expression was up-regulated at 24 h and then recovered to the normal level. In the salinity 49 treatment group, the transcriptional levels of EcproPO mRNA were up-regulated at 24, 72 and 96 h and down-regulated at 48 h (Fig. 4b).
Figure
4.
The expression profiles of the EcproPO gene in adult E. carinicauda after low salinity (a) and high salinity (b) stress. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
3.4
PO activity of juvenile and adult E. carinicauda under low and high salinity stress
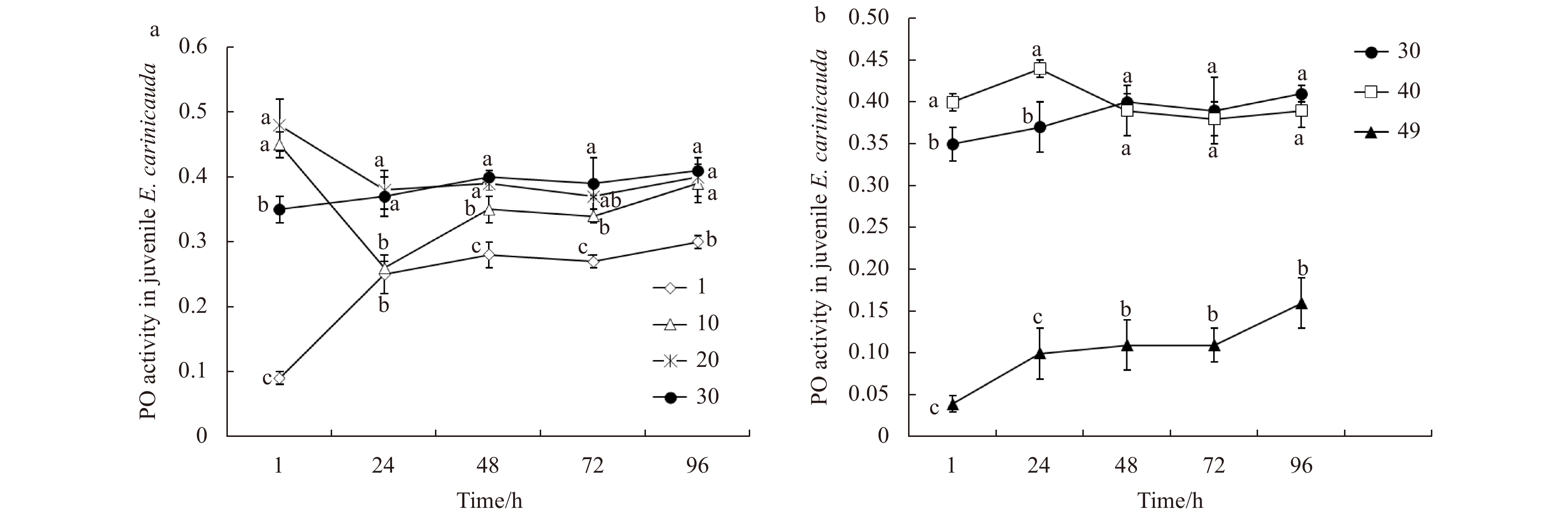
Changes of PO activity of juvenile E. carinicauda under low and high salinity stress are shown in Fig. 5. Compared with the control group (salinity 30), under low salinity stress, the PO activity showed significant decreases after transfer to the extremely low salinity condition (salinity 1). In the salinity 10 treatment group, the maximal reduction of PO activity appeared at 24 h, and then the PO activity increased gradually and recovered at 96 h. No differential PO activity was observed between the salinity 20 and 30 groups after 24 h (Fig. 5a). Under high salinity stress, PO activities in the salinity 40 treatment group in the first 24 h were higher than that in the salinity 30 group and then recovered to the normal level. Shrimp PO activities under extremely high salinity stress (salinity 49) were always lower than those in the salinity 30 group throughout the experiment (Fig. 5b). Phenoloxidase activities in low salinities were higher than high salinities.
Figure
5.
Phenoloxidase (PO) activity of juvenile E. carinicauda after exposure to low salinity (a) and high salinity (b) stress treatments. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
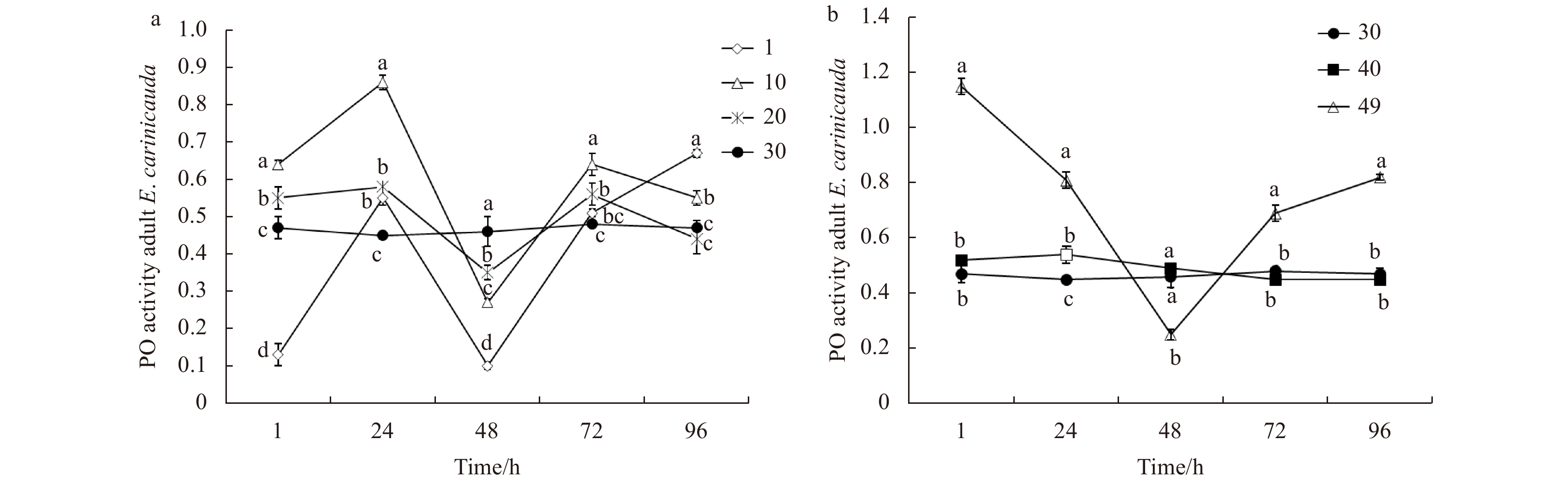
For E. carinicauda adults, changes of PO activity under low and high salinity stress are shown in Fig. 6. Under sudden low salinity stress, the PO activity had a waving trend (Fig. 6a). Under high salinity stress, PO activity in the salinity 40 treatment group showed no obvious difference compared to the salinity 30 group. The PO activities in the salinity 49 treatment group were higher at 1, 24, 72 and 96 h than those in the salinity 30 group (Fig. 6b).
Figure
6.
Phenoloxidase (PO) activity of adult E. carinicauda after exposure low salinity (a) and high salinity (b) stress treatments. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
Acute changes of salinity can significantly effect the survival and immunity of crustaceans, even though they have a greater ability to survive extreme salinity conditions (Le Moullac et al., 1998; Jiravanichpaisal et al., 2004; Li and Chen, 2008; Wang and Chen, 2006; Pan et al., 2010). Previous studies have demonstrated that the optimum salinity range for farming E. carinicauda is between 20–28 (Wang et al., 2005). In this study, E. carinicauda juveniles could survive at a low salinity of 3 and a high salinity of 40 with survival rates more than 95%, indicating a strong tolerance to salinity changes. In addition, the survival rates of E. carinicauda juveniles under low salinity stress were higher than those of high salinity stress, indicating that the E. carinicauda juveniles might have stronger tolerance to low salinity conditions than high salinity conditions. For E. carinicauda adults, survival rates at salinity 1 were significantly less than that at salinity 3. Gu et al. (2004) reported that the survival rate of E. carinicauda was drastically reduced when salinity decreased below 1.08, which was similar to our results. Our preliminary study showed that the salinity tolerance range of adult E. carinicauda was 6–44, which was narrower than that of juveniles.
As a key immune gene, the expression of LGBP has been used to evaluate the effect of salinity stress in crustaceans (Lin et al., 2012). For juvenile E. carinicauda, the initially down-regulated expression levels of LGBP under 1 salinity stress indicated that the immune system might be weakened by extremely low salinity at the onset of salinity stress. Then, the expression levels of LGBP were up-regulated, demonstrating that EcLGBP could be related to the induction of the innate immune system for resisting short term detrimental consequences from low salinity. It is well known that crustaceans are easily vulnerable to diseases under high stresses (Vargas-Albores et al., 1998; Cheng and Chen, 2000). Hence, the down-regulation of EcLGBP mRNA at high salinity conditions implied that immune molecules in the defense system of juveniles were inhibited under high salinity stress. For the adult E. carinicauda, EcLGBP expression was initially up-regulated under all salinity stress treatments, which showed that the induction of this gene could respond to variation of the salinity timely. Further, the fluctuation of the EcLGBP expression levels at 48–96 h might be related to the melanization process and the loss of reproductive capacity (De Gregorio et al., 2002; Pascual et al., 2003).
The proPO system is considered a constituent of the humoral immune system and can be activated by different chemical and several microbial elicitors in crustaceans (Hernández-López et al., 1996). For juvenile E. carinicauda, the expression levels of EcproPO were negatively affected by hyper-salinity, which could lead to a reduction of the immunity of shrimp. The up-regulated expressions of EcproPO under extremely low salinity stress suggested that this gene played an important role in low salinity conditions. The expression tendency of proPO in juveniles in this study was partially in agreement with the preceding observation in gills of E. carinicauda after low salinity stress (4–25) (Li et al., 2015). Previous studies have reported that prophenoloxidase-activating enzyme 1a was up-regulated in L. vannamei in response to low salinity, suggesting that L. vannamei induced its phenoloxidation cascade and cleaved proPO to release PO after low salinity stress (Nikapitiya et al., 2014). For adult E. carinicauda, the significantly up-regulated expression levels of EcproPO under both high and low salinity stress indicated that salinity changes in the environment could induce the immune responses in crustaceans. Therefore, EcproPO could be considered a responsive gene and potential candidate for developing gene markers to evaluate shrimp immunity variations in the environment.
Phenoloxidase is the terminal enzyme in the proPO system that has been proposed as a potential health indicator in shrimp haemolymph (Joseph and Philip, 2007). In the present study, for juvenile E. carinicauda, the PO activity decreased in extreme hypo/hyper-salinity treatments, while initially increased in moderate hypo/hyper-salinity, which indicated that the appropriate hypo/hyper-salinity could increase the immune system, but short-term extremely low/high salinity could weaken immune protection potentially resulting in death. Further, the PO activity values were higher in salinity 20–40 than the values in other salinity treatments, suggesting that juvenile E. carinicauda reared in salinity 20–40 should be the optimal salinity range to maintain the immune system. Moreover, the PO activity values in hypo-salinity conditions were relatively higher than that in hyper-salinity, which indicated that juveniles might have higher tolerance to resistant pathogens in low salinity. An increasing trend in PO activity of Panulirus homarus from salinity 20 to 45 could be due to significant reduction in plasma inhibitors regulating the proPO system (Verghese et al., 2007), which was in agreement with the tendency of PO activity in this study. For adult E. carinicauda, PO activity increased directly under salinity stress in the first 24 h, which might be correlated with the higher resistance of shrimp against pathogen infection by increasing its immunity. White shrimp, L. vannamei, transferred from a salinity of 35 to 25, 20, and 15 exhibited significant decreases in immunity as evidenced by reductions in immune parameters including PO activity after 1–6 h (Li et al., 2010). Hence, adult shrimp farming should be maintained at a constantly high salinity level to prevent exacerbated decreases in innate immunity of shrimp when infected by pathogens, as low-salinity stress led to mortality.
In conclusion, E. carinicauda juveniles could survive in a wider range of salinity conditions than adults. For juvenile E. carinicauda, the expression levels of EcLGBP and EcProPO were up-regulated against low salinities, and PO activities in low salinities were higher compared to those in high salinities. For adult E. carinicauda, the expression profiles of EcLGBP and EcproPO had a different trend of up-regulation under hypo/hyper-salinity stress treatments. The salinity tolerance range of immunity for juvenile and adult E. carinicauda is 20–40 and 30–40, respectively.
Ai Huashui, Huang Yongchun, Li Sedong, et al. 2008. Characterization of a prophenoloxidase from hemocytes of the shrimp Litopenaeus vannamei that is down-regulated by white spot syndrome virus. Fish & Shellfish Immunology, 25(1–2): 28–39
[2]
Amparyup P, Charoensapsri W, Tassanakajon A. 2013. Prophenoloxidase system and its role in shrimp immune responses against major pathogens. Fish & Shellfish Immunology, 34(4): 990–1001
[3]
Ashida M. 1971. Purification and characterization of pre-phenoloxidase from hemolymph of the silkworm Bombyx mori. Archives of Biochemistry and Biophysics, 144(2): 749–762. doi: 10.1016/0003-9861(71)90383-3
[4]
Cerenius L, Lee B L, Söderhäll K. 2008. The proPO-system: pros and cons for its role in invertebrate immunity. Trends in Immunology, 29(6): 263–271. doi: 10.1016/j.it.2008.02.009
[5]
Cerenius L, Söderhäll K. 2004. The prophenoloxidase-activating system in invertebrates. Immunological Reviews, 198(1): 116–126. doi: 10.1111/j.0105-2896.2004.00116.x
[6]
Chen Jinghua, Mai Kangsen, Ma Hongming, et al. 2007. Effects of dissolved oxygen on survival and immune responses of scallop (Chlamys farreri jones et preston). Fish & Shellfish Immunology, 22(3): 272–281
[7]
Cheng W, Chen J C. 2000. Effects of pH, temperature and salinity on immune parameters of the freshwater prawn Macrobrachium rosenbergii. Fish & Shellfish Immunology, 10(4): 387–391
[8]
Cheng W, Liu C H, Tsai C H, et al. 2005. Molecular cloning and characterisation of a pattern recognition molecule, lipopolysaccharide- and β-1, 3-glucan binding protein (LGBP) from the white shrimp Litopenaeus vannamei. Fish & Shellfish Immunology, 18(4): 297–310
[9]
Cieluch U, Charmantier G, Grousset E, et al. 2005. Osmoregulation, immunolocalization of Na+/K+-ATPase, and ultrastructure of branchial epithelia in the developing brown shrimp, Crangon crangon (Decapoda, Caridea). Physiological and Biochemical Zoology, 78(6): 1017–1025. doi: 10.1086/432856
[10]
De Gregorio E, Han S J, Lee W J, et al. 2002. An immune-responsive Serpin regulates the melanization cascade in Drosophila. Developmental Cell, 3(4): 581–592. doi: 10.1016/S1534-5807(02)00267-8
[11]
de la Vega E, Degnan B M, Hall M R, et al. 2007. Differential expression of immune-related genes and transposable elements in black tiger shrimp (Penaeus monodon) exposed to a range of environmental stressors. Fish & Shellfish Immunology, 23(5): 1072–1088
[12]
Dong Cunyou. 1989. The biological properties of Exopalaemon carinicauda in Zhujiang river estuary. Sichuan Journal of Zoology (in Chinese), 8(4): 36–38
[13]
Duan Yafei, Liu Ping, Li Jitao, et al. 2013. cDNA cloning, characterization and expression analysis of peroxiredoxin 5 gene in the ridgetail white prawn Exopalaemon carinicauda. Molecular Biology Reports, 40(12): 6569–6577. doi: 10.1007/s11033-013-2702-4
[14]
Gu Jun, Li Guofeng, Zhang Zhenhua, et al. 2004. The response of Exopalaemon carinicauda to acute salinity stress. Journal of Aquaculture (in Chinese), 25(2): 39–40
[15]
Hernández-López J, Gollas-Galván T, Vargas-Albores F. 1996. Activation of the prophenoloxidase system of the brown shrimp (Penaeus californiensis Holmes). Comparative Biochemistry and Physiology Part C: Pharmacology, Toxicology and Endocrinology, 113(1): 61–66. doi: 10.1016/0742-8413(95)02033-0
[16]
Hu Fawen. 2008. Effect of dopamine injection, salinity changes on the prophenoloxidase system and defence parameters of the White Shrimp Litopenaeus vannamei (in Chinese) [dissertation]. Qingdao: Ocean University of China
[17]
Ishwarya R, Jayanthi S, Muthulakshmi P, et al. 2016. Immune indices and identical functions of two prophenoloxidases from the haemolymph of green tiger shrimp Penaeus semisulcatus and its antibiofilm activity. Fish & Shellfish Immunology, 51: 220–228
[18]
Jiravanichpaisal P, Puanglarp N, Petkon S, et al. 2007. Expression of immune-related genes in larval stages of the giant tiger shrimp, Penaeus monodon. Fish Shellfish Immunol, 23(4): 815–824. doi: 10.1016/j.fsi.2007.03.003
[19]
Jiravanichpaisal P, Söderhäll K, Söderhäll I. 2004. Effect of water temperature on the immune response and infectivity pattern of white spot syndrome virus (WSSV) in freshwater crayfish. Fish & Shellfish Immunology, 17(3): 265–275
[20]
Johansson M W, Soderhall K. 1989. Cellular immunity in crustaceans and the proPO system. Parasitology Today, 5(6): 171–176. doi: 10.1016/0169-4758(89)90139-7
[21]
Joseph A, Philip R. 2007. Acute salinity stress alters the haemolymph metabolic profile of Penaeus monodon and reduces immunocompetence to white spot syndrome virus infection. Aquaculture, 272(1–4): 87–97. doi: 10.1016/j.aquaculture.2007.08.047
[22]
Kumlu M, Eroldogan O T, Aktas M. 2000. Effects of temperature and salinity on larval growth, survival and development of Penaeus semisulcatus. Aquaculture, 188(1–2): 167–173. doi: 10.1016/S0044-8486(00)00330-6
[23]
Le Moullac G, Soyez C, Saulnier D, et al. 1998. Effect of hypoxic stress on the immune response and the resistance to vibriosis of the shrimp Penaeus stylirostris. Fish & Shellfish Immunology, 8(8): 621–629
[24]
Lee S Y, Wang Ruigong, Söderhäll K. 2000. A lipopolysaccharide- and β-1, 3-glucan-binding protein from hemocytes of the freshwater crayfish Pacifastacus leniusculus Purification, characterization, and cDNA cloning. Journal of Biological Chemistry, 275(2): 1337–1343. doi: 10.1074/jbc.275.2.1337
[25]
Lei Zhiwen, Huang Jie, Yang Bing, et al. 2001. Immune factors in haemolymph supernatant of Penaeus chinensis infected by WSSV. Journal of Fishery Sciences of China (in Chinese), 8(4): 46–51
[26]
Li Changche, Chen J C. 2008. The immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus under low and high pH stress. Fish & Shellfish Immunology, 25(6): 701–709
[27]
Li Jitao, Han Junying, Chen Ping, et al. 2012. Cloning of a heat shock protein 90(HSP90) gene and expression analysis in the ridgetail white prawn Exopalaemon carinicauda. Fish & Shellfish Immunology, 32(6): 1191–1197
[28]
Li Jitao, Ma Peng, Liu Ping, et al. 2015. The roles of Na+/K+-ATPase α-subunit gene from the ridgetail white prawn Exopalaemon carinicauda in response to salinity stresses. Fish & Shellfish Immunology, 42(2): 264–271
[29]
Li Fuhua, Xiang Jianhai. 2013. Recent advances in researches on the innate immunity of shrimp in China. Developmental & Comparative Immunology, 39(1–2): 11–26
[30]
Li Changche, Yeh S T, Chen J C. 2010. Innate immunity of the white shrimp Litopenaeus vannamei weakened by the combination of a Vibrio alginolyticus injection and low-salinity stress. Fish & Shellfish Immunology, 28(1): 121–127
[31]
Lin Yong-Chin, Chen Jiann-Chu, Li Changche, et al. 2012. Modulation of the innate immune system in white shrimp Litopenaeus vannamei following long-term low salinity exposure. Fish & Shellfish Immunology, 33(2): 324–331
[32]
Liu Fengsong, Li Fuhua, Dong Bo, et al. 2009. Molecular cloning and characterisation of a pattern recognition protein, lipopolysaccharide and β-1, 3-glucan binding protein (LGBP) from Chinese shrimp Fenneropenaeus chinensis. Molecular Biology Reports, 36(3): 471–477. doi: 10.1007/s11033-007-9203-2
[33]
Livak K J, Schmittgen T D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔ CT method. Methods, 25(4): 402–408. doi: 10.1006/meth.2001.1262
[34]
Nikapitiya C, Kim W S, Park K, et al. 2014. Identification of potential markers and sensitive tissues for low or high salinity stress in an intertidal mud crab (Macrophthalmus japonicus). Fish & Shellfish Immunology, 41(2): 407–416
[35]
Pan Luqing, Xie Peng, Hu Fawen. 2010. Responses of prophenoloxidase system and related defence parameters of Litopenaeus vannamei to low salinity. Journal of Ocean University of China, 9(3): 273–278. doi: 10.1007/s11802-010-1711-3
[36]
Pan Luqing, Zhang Linjuan, Liu Hongyu. 2007. Effects of salinity and pH on ion-transport enzyme activities, survival and growth of Litopenaeus vannamei postlarvae. Aquaculture, 273(4): 711–720. doi: 10.1016/j.aquaculture.2007.07.218
[37]
Pascual C, Sánchez A, Sánchez A, et al. 2003. Haemolymph metabolic variables and immune response in Litopenaeus setiferus adult males: the effect of an extreme temperature. Aquaculture, 218(1–4): 637–650. doi: 10.1016/S0044-8486(02)00300-9
[38]
Söderhäll K, Cerenius L. 1992. Crustacean immunity. Annual Review of Fish Diseases, 2: 3–23. doi: 10.1016/0959-8030(92)90053-Z
[39]
Söderhäll K, Smith V J. 1983. Separation of the haemocyte populations of Carcinus maenas and other marine decapods. Developmental & Comparative Immunology, 7(2): 229–239
[40]
Vargas-Albores F, Hinojosa-Baltazar P, Portillo-Clark G, et al. 1998. Influence of temperature and salinity on the yellowleg shrimp, Penaeus californieinsis Holmes, prophenoloxidase system. Aquaculture Research, 29(8): 549–553. doi: 10.1046/j.1365-2109.1998.00235.x
[41]
Verghese B, Radhakrishnan E V, Padhi A. 2007. Effect of environmental parameters on immune response of the Indian spiny lobster, Panulirus homarus (Linnaeus, 1758). Fish & Shellfish Immunology, 23(5): 928–936
[42]
Wang F I, Chen J C. 2006. Effect of salinity on the immune response of tiger shrimp Penaeus monodon and its susceptibility to Photobacterium damselae subsp. Damselae. Fish & Shellfish Immunology, 20(5): 671–681
[43]
Wang Xingqiang, Sudha K, Mei Cao, et al. 2010. Effects of low salinity and low temperature on survival, growth, and energy budget of juvenile Exopalaemon carinicauda. Journal of Shellfish Research, 29(4): 1035–1041. doi: 10.2983/035.029.0405
[44]
Wang Xingqiang, Yan Binlun, Ma Shen, et al. 2005. Study on the biology and cultural ecology of Exopalaemon carinicauda. Shandong Fisheries, 22(8): 21–23
[45]
Xu Wenjun, Xie Jianjun, Shi Hui, et al. 2010. Hematodinium infections in cultured ridgetail white prawns, Exopalaemon carinicauda, in eastern China. Aquaculture, 300(1–4): 25–31. doi: 10.1016/j.aquaculture.2009.12.024
[46]
Yang Ya’nan, Bao Chenchang, Liu An, et al. 2014. Immune responses of prophenoloxidase in the mud crab Scylla paramamosain against Vibrio alginolyticus infection: in vivo and in vitro gene silencing evidence. Fish & Shellfish Immunology, 39(2): 237–244
[47]
Zhang Chengsong, Li Fuhua, Xiang Jianhai. 2014. Effect of salinity on growth and first sexual maturity of Exopalaemon carinicauda (Holthuis, 1950). Chinese Journal of Oceanology and Limnology, 32(1): 65–70. doi: 10.1007/s00343-014-3040-7
[48]
Zhao Qun, Pan Luqing, Ren Qin, et al. 2015. Digital gene expression analysis in hemocytes of the white shrimp Litopenaeus vannamei in response to low salinity stress. Fish & Shellfish Immunology, 42(2): 400–407
Rongxiao Wang, Yuke Bu, Kefan Xing, et al. Integrated analysis of transcriptome and metabolome reveals chronic low salinity stress responses in the muscle of Exopalaemon carinicauda. Comparative Biochemistry and Physiology Part D: Genomics and Proteomics, 2024, 52: 101340. doi:10.1016/j.cbd.2024.101340
2.
Jiajia Wang, Jianjian Lv, Miao Shi, et al. Chromosome-level genome assembly of ridgetail white shrimp Exopalaemon carinicauda. Scientific Data, 2024, 11(1) doi:10.1038/s41597-024-03423-9
3.
Wenjun Shi, Runhao Hu, Ran Zhao, et al. Transcriptome analysis of hepatopancreas and gills of Palaemon gravieri under salinity stress. Gene, 2023, 851: 147013. doi:10.1016/j.gene.2022.147013
4.
Reyhaneh Ravanbakhsh, Naser Agh, Mojtaba Nouraein, et al. Prolonged ecological changes can affect morphometrics and gene expression profile? Focusing on Hsp-70 and NLHS-induced Hsp-70 of Artemia urmiana. Environmental Research, 2023, 238: 117254. doi:10.1016/j.envres.2023.117254
5.
Saravanan Rajendran, Sugumar Vasudevan. Activation of prophenoloxidase and hyperglycemia as indicators of microbial stress in the blue swimmer crab Portunus pelagicus. Marine Pollution Bulletin, 2020, 160: 111711. doi:10.1016/j.marpolbul.2020.111711
Figure 1. The expression profiles of the EcLGBP gene in juvenile E. carinicauda after low salinity (a) and high salinity (b) stress. Vertical bars represent the mean±SE (n=3). Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
Figure 2. The expression profiles of the EcLGBP gene in adult E. carinicauda after low salinity (a) and high salinity (b) stress. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
Figure 3. The expression profiles of the EcproPO gene in juvenile E. carinicauda after exposure to low salinity (a) and high salinity (b) stress. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
Figure 4. The expression profiles of the EcproPO gene in adult E. carinicauda after low salinity (a) and high salinity (b) stress. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
Figure 5. Phenoloxidase (PO) activity of juvenile E. carinicauda after exposure to low salinity (a) and high salinity (b) stress treatments. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.
Figure 6. Phenoloxidase (PO) activity of adult E. carinicauda after exposure low salinity (a) and high salinity (b) stress treatments. Data without shared letters were significantly different (P<0.05) among treatments at the same exposure time.



 DownLoad:
DownLoad:

 DownLoad:
DownLoad:










 DownLoad:
DownLoad: