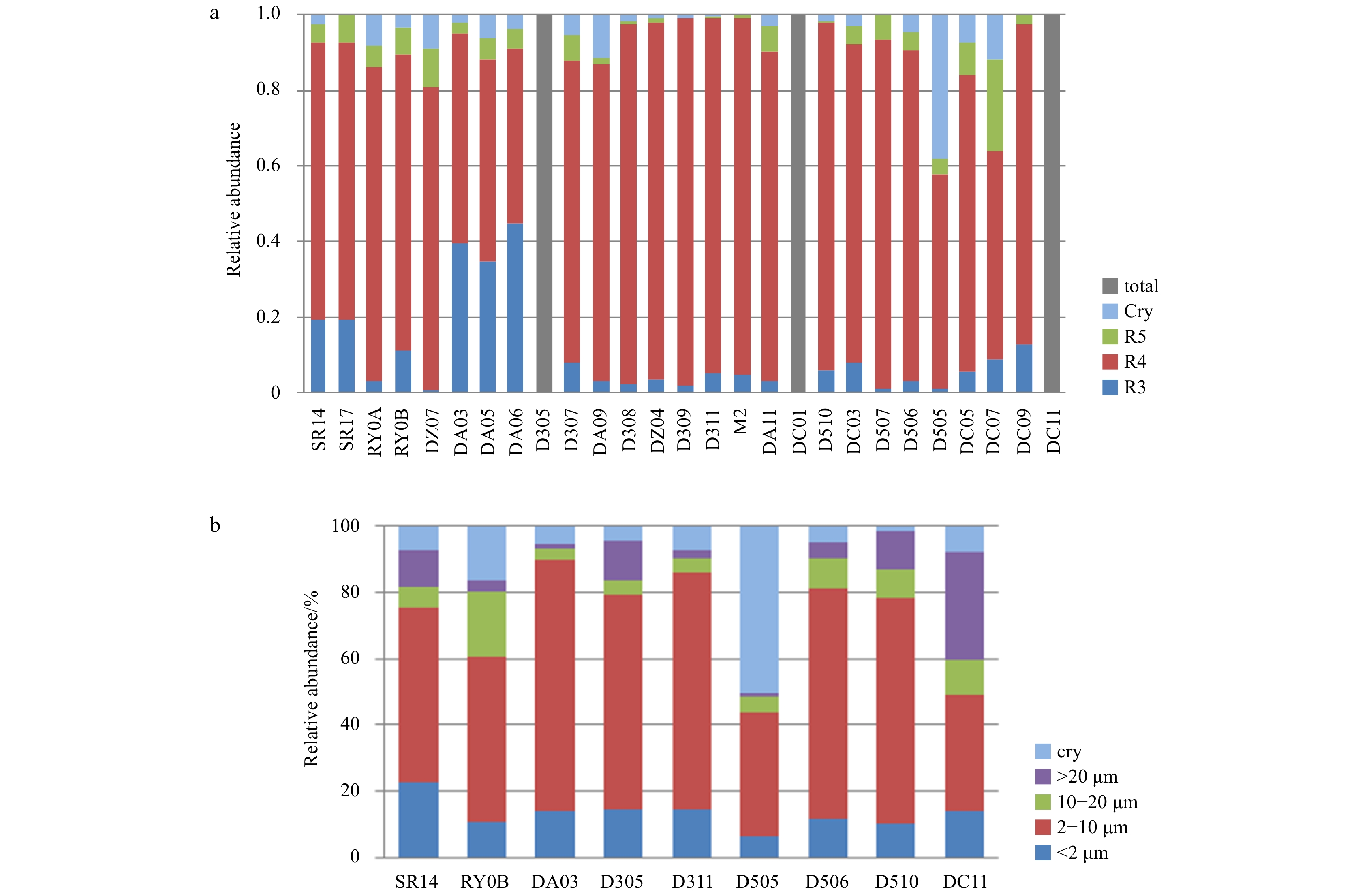
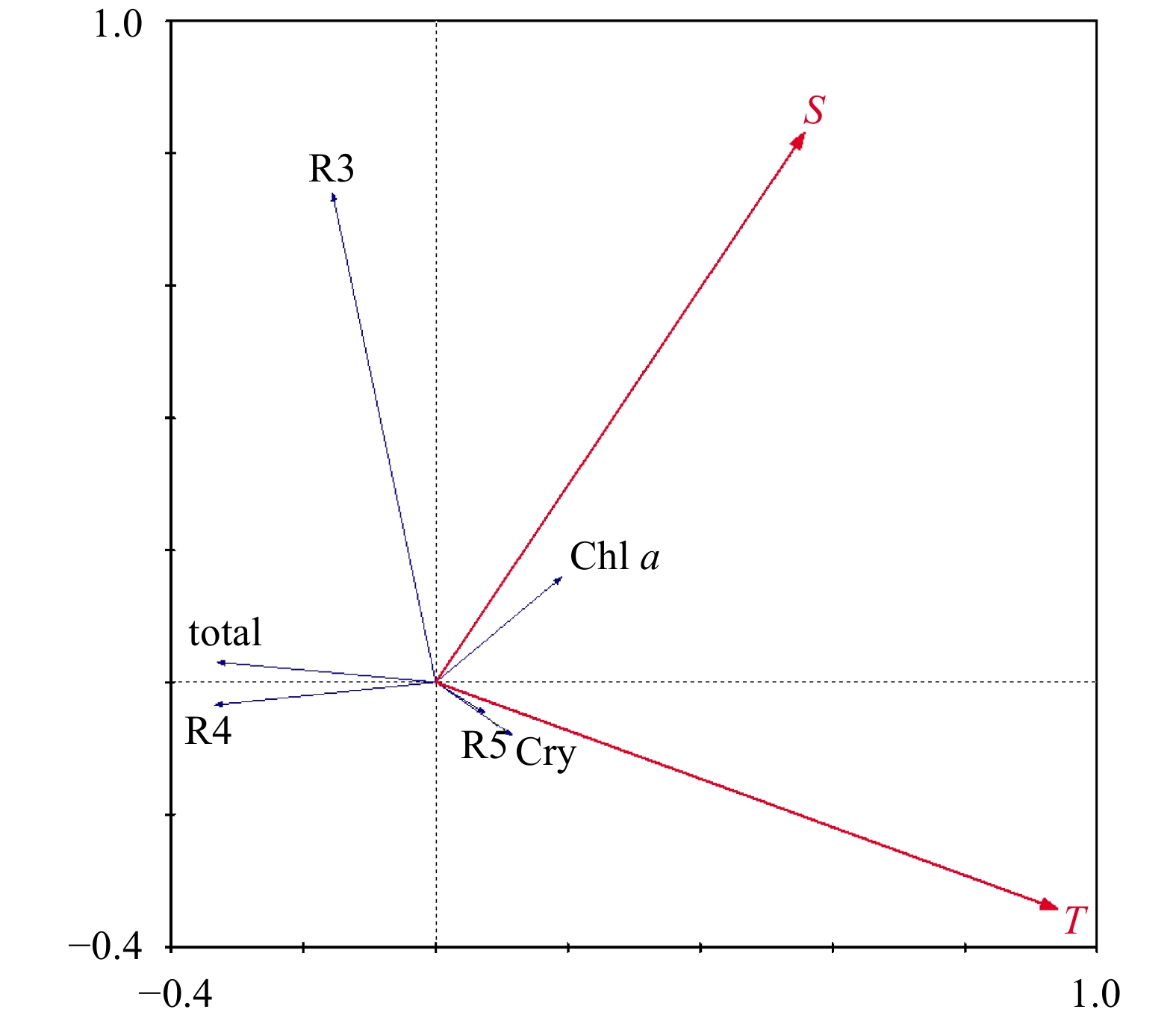
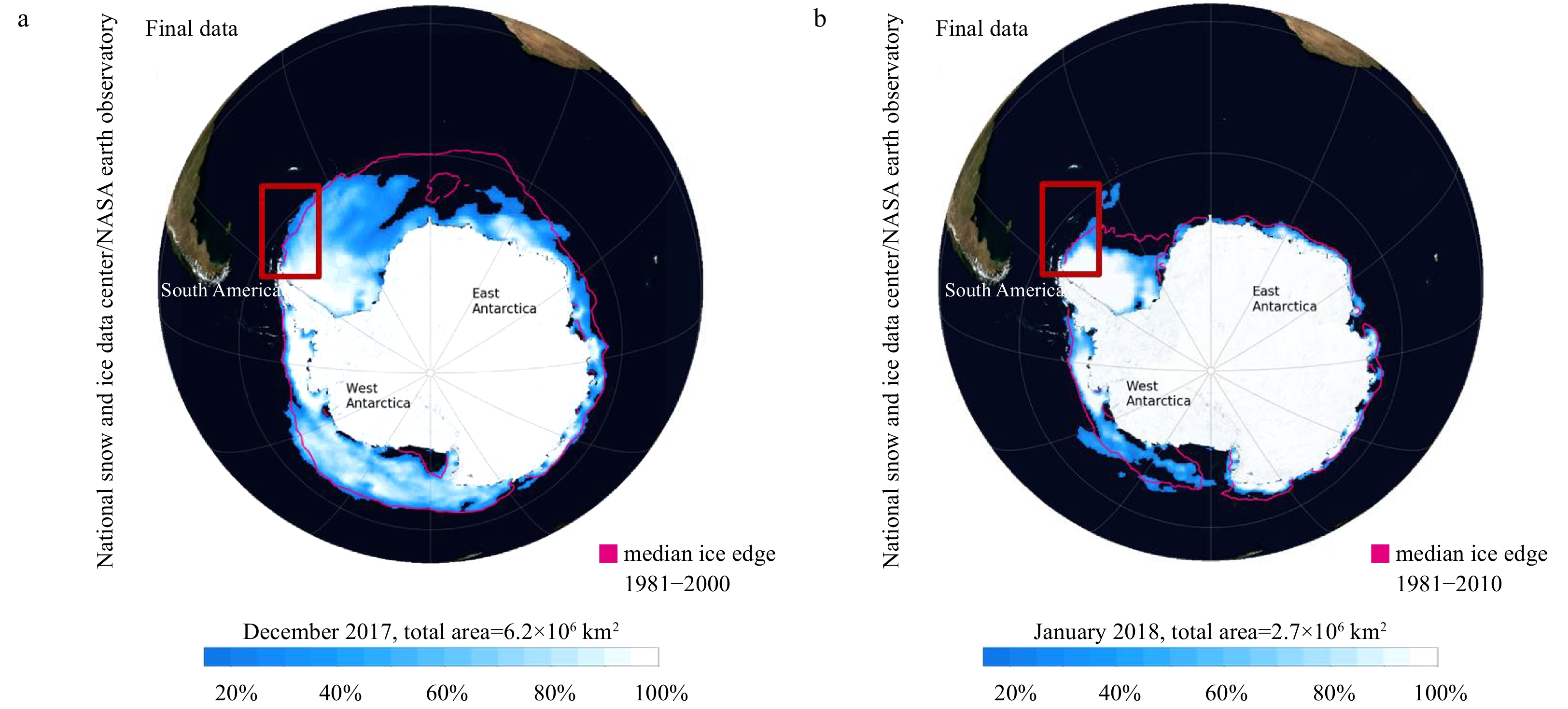
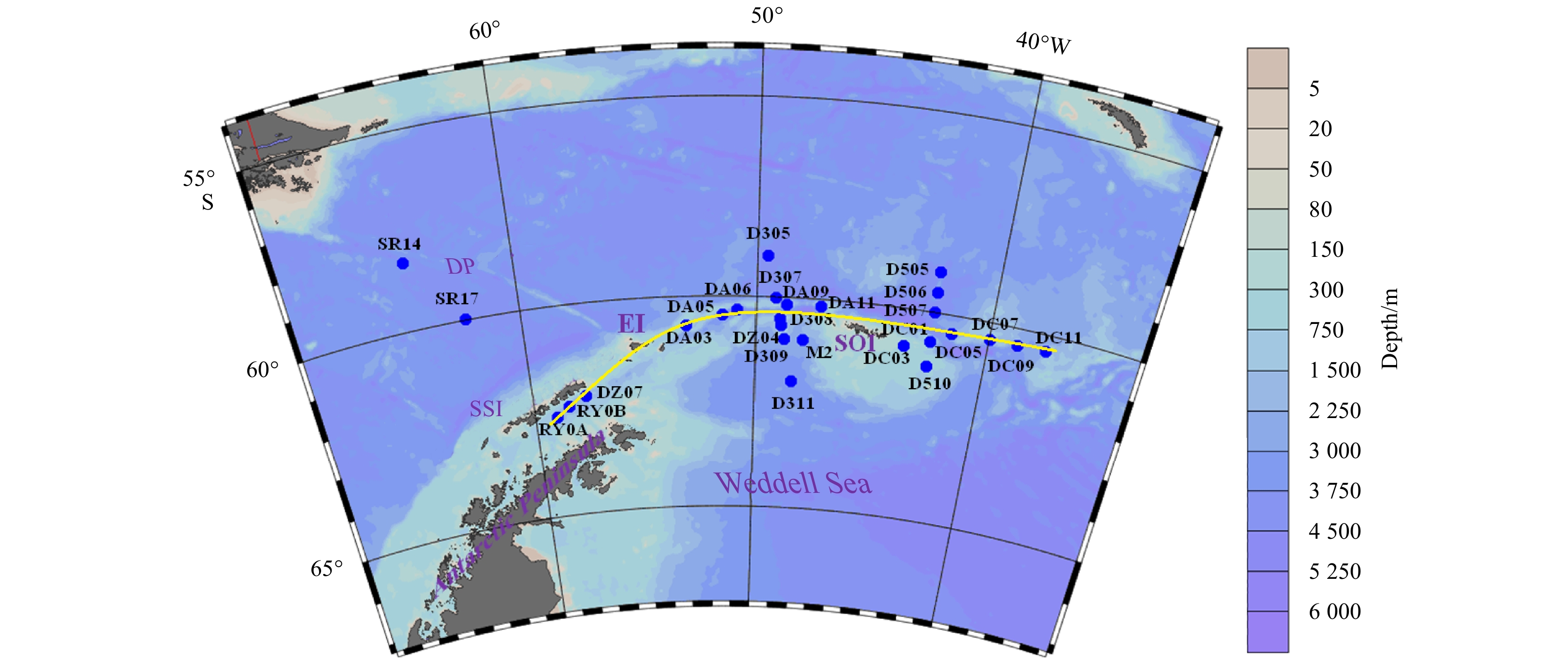
| Citation: | Lu Liu, Mingzhu Fu, Kaiming Sun, Qinzeng Xu, Zongjun Xu, Xuelei Zhang, Zongling Wang. The distribution of phytoplankton size and major influencing factors in the surface waters near the northern end of the Antarctic Peninsula[J]. Acta Oceanologica Sinica, 2021, 40(6): 92-99. doi: 10.1007/s13131-020-1611-3

|
| [1] |
Agawin N S R, Agustí S, Duarte C M. 2002. Abundance of Antarctic picophytoplankton and their response to light and nutrient manipulation. Aquatic Microbial Ecology, 29(2): 161–172
|
| [2] |
Becquevort S. 1997. Nanoprotozooplankton in the Atlantic sector of the Southern Ocean during early spring: biomass and feeding activities. Deep-Sea Research Part II: Topical Studies in Oceanography, 44(1–2): 355–373. doi: 10.1016/S0967-0645(96)00076-8
|
| [3] |
Blanchot J, Rodier M. 1996. Picophytoplankton abundance and biomass in the western tropical Pacific Ocean during the 1992 El Niño year: results from flow cytometry. Deep-Sea Research Part I: Oceanographic Research Papers, 43(6): 877–895. doi: 10.1016/0967-0637(96)00026-X
|
| [4] |
Buma A G J, Gieskes W W C, Thomsen H A. 1992. Abundance of Cryptophyceae and chlorophyll b-containing organisms in the Weddell-Scotia Confluence area in the spring of 1988. Polar Biology, 12(1): 43–52
|
| [5] |
Cermeño P, Marañón E, Rodríguez J, et al. 2005. Size dependence of coastal phytoplankton photosynthesis under vertical mixing conditions. Journal of Plankton Research, 27(5): 473–483. doi: 10.1093/plankt/fbi021
|
| [6] |
Egas C, Henríquez-Castillo C, Delherbe N, et al. 2017. Short timescale dynamics of phytoplankton in Fildes Bay, Antarctica. Antarctic Science, 29(3): 217–228. doi: 10.1017/S0954102016000699
|
| [7] |
Fetterer F, Knowles K, Meier W N, et al. 2017 Updated Daily. Sea Ice Index, Version 3. [Antarctic, Sea Ice Concentration]. Boulder, Colorado USA: National Snow and Ice Data Center
|
| [8] |
García-Muñoz C, Lubián L M, García C M, et al. 2013. A mesoscale study of phytoplankton assemblages around the South Shetland Islands (Antarctica). Polar Biology, 36(8): 1107–1123. doi: 10.1007/s00300-013-1333-5
|
| [9] |
García-Muñoz C, Sobrino C, Lubián L M, et al. 2014. Factors controlling phytoplankton physiological state around the South Shetland Islands (Antarctica). Marine Ecology Progress Series, 498: 55–71. doi: 10.3354/meps10616
|
| [10] |
Garibotti I A, Vernet M, Ferrario M E, et al. 2003. Phytoplankton spatial distribution patterns along the western Antarctic Peninsula (Southern Ocean). Marine Ecology Progress Series, 261: 21–39. doi: 10.3354/meps261021
|
| [11] |
Garibotti I A, Vernet M, Ferrario M E. 2005. Annually recurrent phytoplanktonic assemblages during summer in the seasonal ice zone west of the Antarctic Peninsula (Southern Ocean). Deep-Sea Research Part I: Oceanographic Research Papers, 52(10): 1823–1841. doi: 10.1016/j.dsr.2005.05.003
|
| [12] |
Green R E, Sosik H M, Olson R J. 2003. Contributions of phytoplankton and other particles to inherent optical properties in New England continental shelf waters. Limnology and Oceanography, 48(6): 2377–2391. doi: 10.4319/lo.2003.48.6.2377
|
| [13] |
Hewes C D. 2009. Cell size of Antarctic phytoplankton as a biogeochemical condition. Antarctic Science, 21(5): 457–470. doi: 10.1017/S0954102009990125
|
| [14] |
Hewes C D, Holm-Hansen O, Sakshaug E. 1985. Alternate carbon pathways at lower trophic levels in the Antarctic food web. In: Siegfried W R, Condy P R, Laws R M, eds. Antarctic Nutrient Cycles and Food Webs. Berlin, Heidelberg: Springer, 277–283
|
| [15] |
Holm-Hansen O, Lorenzen C J, Holmes R W, et al. 1965. Fluorometric determination of chlorophyll. ICES Journal of Marine Science, 30(1): 3–15. doi: 10.1093/icesjms/30.1.3
|
| [16] |
Ishikawa A, Wright S W, Van Den Enden R, et al. 2002. Abundance, size structure and community composition of phytoplankton in the Southern Ocean in the austral summer 1999/2000. Polar Bioscience, 15: 11–26
|
| [17] |
Jacques G, Panouse M. 1991. Biomass and composition of size fractionated phytoplankton in the Weddell-Scotia Confluence area. Polar Biology, 11(5): 315–328
|
| [18] |
Korb R E, Whitehouse M J, Gordon M, et al. 2010. Summer microplankton community structure across the Scotia Sea: implications for biological carbon export. Biogeosciences, 7(1): 343–356. doi: 10.5194/bg-7-343-2010
|
| [19] |
Lepš J, Šmilauer P. 2003. Multivariate Analysis of Ecological Data Using CANOCO. Cambridge: Cambridge University Press
|
| [20] |
Marañón E. 2015. Cell Size as a key determinant of phytoplankton metabolism and community structure. Annual Review of Marine Science, 7: 241–264. doi: 10.1146/annurev-marine-010814-015955
|
| [21] |
Marañón E, Cermeño P, Latasa M, et al. 2012. Temperature, resources, and phytoplankton size structure in the ocean. Limnology and Oceanography, 57(5): 1266–1278. doi: 10.4319/lo.2012.57.5.1266
|
| [22] |
Morán X A G, López-Urrutia Á, Calvo-Díaz A, et al. 2010. Increasing importance of small phytoplankton in a warmer ocean. Global Change Biology, 16: 1137-44.
|
| [23] |
Olson R J, Sosik H M, Chekalyuk A M, et al. 2000. Effects of iron enrichment on phytoplankton in the Southern Ocean during late summer: active fluorescence and flow cytometric analyses. Deep-Sea Research Part II: Topical Studies in Oceanography, 47(15–16): 3181–3200. doi: 10.1016/S0967-0645(00)00064-3
|
| [24] |
Rodríguez J, Jiménez-Gómez F, Blanco J M, et al. 2002. Physical gradients and spatial variability of the size structure and composition of phytoplankton in the Gerlache Strait (Antarctica). Deep-Sea Research Part II: Topical Studies in Oceanography, 49(4–5): 693–706. doi: 10.1016/S0967-0645(01)00119-9
|
| [25] |
Sakshaug E, Holm-Hansen O. 1986. Photoadaptation in Antarctic phytopfankton: variations in growth rate, chemical composition and P versus I curves. Journal of Plankton Research, 8(3): 459–473. doi: 10.1093/plankt/8.3.459
|
| [26] |
Schloss I, Estrada M. 1994. Phytoplankton composition in the Weddell-Scotia Confluence area during austral spring in relation to hydrography. Polar Biology, 14(2): 77–90
|
| [27] |
Smith R C, Martinson D G, Stammerjohn S E, et al. 2008. Bellingshausen and western Antarctic Peninsula region: pigment biomass and sea-ice spatial/temporal distributions and interannual variabilty. Deep-Sea Research Part II: Topical Studies in Oceanography, 55(18–19): 1949–1963. doi: 10.1016/j.dsr2.2008.04.027
|
| [28] |
Sosik H M, Olson J J, Armbrust E V. 2010. Flow cytometry in phytoplankton research. In: Suggett D J, Prášil O, Borowitzka M A, eds. Chlorophyll a Fluorescence in Aquatic Sciences: Methods and Applications. Dordrecht: Springer, 171–185
|
| [29] |
Vernet M, Martinson D, Iannuzzi R, et al. 2008. Primary production within the sea-ice zone west of the Antarctic Peninsula: I. Sea ice, summer mixed layer, and irradiance. Deep-Sea Research Part II: Topical Studies in Oceanography, 55(18–19): 2068–2085. doi: 10.1016/j.dsr2.2008.05.021
|
| [30] |
Ward B A, Dutkiewicz S, Follows M J. 2014. Modelling spatial and temporal patterns in size-structured marine plankton communities: top–down and bottom–up controls. Journal of Plankton Research, 36(1): 31–47. doi: 10.1093/plankt/fbt097
|
| [31] |
Zhang Fang, Ma Yuxin, Lin Ling, et al. 2012. Hydrophysical correlation and water mass indication of optical physiological parameters of picophytoplankton in Prydz Bay during autumn 2008. Journal of Microbiological Methods, 91(3): 559–565. doi: 10.1016/j.mimet.2012.09.030
|

Figures(8)

Supported by:
Beijing Renhe Information Technology Co. Ltd

Lu Liu, Mingzhu Fu, Kaiming Sun, Qinzeng Xu, Zongjun Xu, Xuelei Zhang, Zongling Wang. The distribution of phytoplankton size and major influencing factors in the surface waters near the northern end of the Antarctic Peninsula[J]. Acta Oceanologica Sinica, 2021, 40(6): 92-99. doi: 10.1007/s13131-020-1611-3




 DownLoad:
DownLoad: