Yuanyuan Ding, Yao Bian, Huina Wang, Jing Liu, Jingrui Li, Hongwei Wang. Early development, life history and ecological habits of Grateloupia constricata Li et Ding[J]. Acta Oceanologica Sinica, 2020, 39(10): 155-161. doi: 10.1007/s13131-020-1662-5
Citation:
Yuanyuan Ding, Yao Bian, Huina Wang, Jing Liu, Jingrui Li, Hongwei Wang. Early development, life history and ecological habits of Grateloupia constricata Li et Ding[J]. Acta Oceanologica Sinica, 2020, 39(10): 155-161. doi: 10.1007/s13131-020-1662-5
Yuanyuan Ding, Yao Bian, Huina Wang, Jing Liu, Jingrui Li, Hongwei Wang. Early development, life history and ecological habits of Grateloupia constricata Li et Ding[J]. Acta Oceanologica Sinica, 2020, 39(10): 155-161. doi: 10.1007/s13131-020-1662-5
Citation:
Yuanyuan Ding, Yao Bian, Huina Wang, Jing Liu, Jingrui Li, Hongwei Wang. Early development, life history and ecological habits of Grateloupia constricata Li et Ding[J]. Acta Oceanologica Sinica, 2020, 39(10): 155-161. doi: 10.1007/s13131-020-1662-5
As the largest genus of Halymeniaceae, Grateloupia has been widely reported. Here, we observed the life history and early development of Grateloupia constricata Li et Ding and investigated the effects of temperature, irradiance, and photoperiod on the discoid crust and sporeling development of G. constricata under laboratory conditions. We observed that the type of carpospore development was “mediate discal type”. The life history included homotypic gametophyte (haploid), carposporophyte (diploid), and tetrasporophyte (diploid), with typical isomorphic alternation of generations. The results of double factorial analysis showed that both single factorial effects and interaction among temperature, photoperiod, and irradiance were obviously significant on the discoid crust and sporeling development. Furthermore, we found that the optimum combination of condition for the early growth and development of G. constricata was temperature 20°C, irradiance 80 μmol photons/(m2·s) and photoperiod 16L:8D. This study provides the theoretical basis and technical support for the conservation of the Grateloupia germplasm, artificial breeding, large-scale cultivation and sustainable development.
The marine alga Grateloupiaconstricata (Halymeniaceae, Rhodophyta) was identified as a new species based on the morphological and reproductive structure characteristics of female gametophytes collected by Weixin Li from Qingdao, Shandong Province, China in June 1984 (Xia et al., 2004).
The morphology of G. constricata is distinctive. The thalli are upright, clustered, purplish red, up to 5–10 cm high and 1–1.5 cm wide with pinnately branched 1–2 orders. Abundant branches are opposite, alternate or partial with stick or long cones, usually constricted or tapered at the base. There is a disciform holdfast at the bottom of the algae for adsorbing on rock or shellfish (Fig. 1). As a typical intertidal alga, G. constricata grows on the rocks in the high and medium tidal zones and matures in August (Xia et al., 2004).
Figure
1.
The field habitat of Grateloupia constricata Li et Ding. a. The female gametophyte of G. constricata Li et Ding (arrow), and b. the male gametophyte of G. constricata Li et Ding (arrow).
Grateloupia constricata is an important economic species which has not been artificially cultivated, probably because the life history and ecological habit are still unknown (Fang et al., 2011). In this study, we observed the life history, especially the early development of G. constricata in detail under laboratory condition. We also conducted the double-factorial experiment to investigate the interaction among temperature, irradiance and photoperiod on the discoid crust and sporeling development. We found out the optimum conditions for early growth of G. constricata, which could be the theoretical basis and technical support for the conservation of the Grateloupia germplasm, artificial breeding, large-scale cultivation, sustainable development and utilization.
2.
Materials and methods
2.1
Sample collection and processing
The mature G. constricata (LNU17060506, LNU17060512, LNU17060517, LNU17060565) used in this experiment were collected at the First Swimming Beach, Luxun Park and Maidao of Qingdao, Shandong Province in June 2017. We collected healthy and fully developed gametophyte with mature cystocarps and the tetrasporophyte with tetrasporangia, placed them in an incubator filled with seawater. Female gametophytes with 80% mature cystocarps were chosen and rinsed repeatedly with sterile seawater, and then washed with a brush to remove dirt, micro-organisms and other miscellaneous algae on the surface of the algae. The prepared algae were placed in a well-ventilated area to dry for 1 h, then transferred to a sterile Petri dish with sufficient Provasoli’s Enriched Seawater (PES). Seven glass slides were placed at the bottom in each dish for the spore adhering and collection (Provasoli, 1968). When the number of spores observed with an optical microscope (100×) reached 20–30, the algae were removed. GeO2 (2 mg/L) was added to prevent the growth of other algae and the culture solution was replaced every 2 d. After the straight stereo appeared, the slides were transferred to a culture cylinder (10 cm×10 cm×5 cm) for further cultivation.
2.2
Preculture for experimental condition setting
The spore-attached slides were pre-cultured in an illumination incubator (LHR-250-GB) under the following conditions: temperature (20±2)°C, irradiance 80 μmol photons/(m2·s), salinity 30±1 and photoperiod 12L:12D. When the diameter of discoid crusts reached about 50 μm, the slides were transferred to the specific environmental conditions for further experiments.
2.3
Double factorial experiments
We signed two types of double factorial experiments: temperature vs. irradiance and temperature vs. photoperiod. All experimental factors were remained the same as the preculture conditions except for the double factors we chose. The pre-cultured discoid crusts were placed at temperature of 5°C, 10°C, 15°C, 20°C, 25°C, 30°C, respectively. The irradiances were set to 50, 80 and 110 μmol photons/(m2·s), respectively. The photoperiods were set to 8L:16D, 12L:12D and 16L:8D, respectively. A total of 18 combinations were detected with three parallel samples for each combination (Table 1). We randomly selected ten disks from each experimental group to measure their diameters after 14 d of culture. Sporelings with a height of about 500 μm were placed at the same experimental conditions to measure their heights after 21 d of cultivation.
Table
1.
Combinations of conditions for double factorial experiments
Temperature vs. Irradiance
Temperature vs. Photoperiod
Group
T
I
Group
T
P
1
5
50
1
5
8:16
2
5
80
2
5
12:12
3
5
110
3
5
16:8
4
10
50
4
10
8:16
5
10
80
5
10
12:12
6
10
110
6
10
16:8
7
15
50
7
15
8:16
8
15
80
8
15
12:12
9
15
110
9
15
16:8
10
20
50
10
20
8:16
11
20
80
11
20
12:12
12
20
110
12
20
16:8
13
25
50
13
25
8:16
14
25
80
14
25
12:12
15
25
110
15
25
16:8
16
30
50
16
30
8:16
17
30
80
17
30
12:12
18
30
110
18
30
16:8
Note: T represents temperature (°C), I irradiance (μmol photons/(m2·s)), and P photoperiod (L:D).
Photographs were taken using an Olympus BH2 microscope (Olympus Beijing Co. Ltd., China). The images were mounted on a Nikon camera DL C300-L (Nikon Corporation, Japan) and visualized in Photoshop Adobe System. The progress of protoplast transform was recorded with an Olympus BX53 fluorescence microscope and digitally photographed. The images were captured using the Olympus IPP software package and visualized in Photoshop Adobe System. The relative growth rate (RGR, %/d) of discoid crusts and sporelings were determined using the formula RGR=(lnIt–lnIo)/t×100%. Io represents the initial diameter of discoid crusts or the initial height of sporelings. It represents the discoid crust diameter or sporeling height after t d of cultivation. Double factorial analysis was performed through Two-way ANOVA using SPSS 22.0 according to an adjusted P-value (<0.05). An F-test was used to test for differences in the ranges of SD.
3.
Results
3.1
Early development process of carpospores
Carposporophytes were formed within the female gametophyte cortex of G. constricata. Matured carpospores were crimson and raised on the surface of female gametophyte with diameter of 200–350 μm. Cystocarp holes, with diameter of 50–80 μm, were discovered on the carpospores (Fig. 2a). Afterwards, carpospores overflowed from carpospore holes and attached to the slide (Fig. 2b). The carpospores were spherical or elliptical, light red in color, with diameter of about 15 μm (Fig. 2c). After cultured for 24 h, carpospores began to germinate.
Figure
2.
The development of carpospores in G. constricata Li et Ding. a. Distribution of carposporophytes (ch represents cystocarp hole), b–c. carpospores, d. transformation of protoplasts, e–h. division of carpospores, i. formation of discoid crust, i–k. fusion of discoid crusts, l. formation of upright thalli, m. upright branches, and n–o. growth of sporeling.
The germination progress was as follows: Firstly, protoplast of the carpospores was depressed to one side and moved to the depressed side (Fig. 3a). Meanwhile, the opposite side of the carpospores began to protrude and elongate to form a germination tube. Then, protoplasts began to move towards to the germination tubes from the carpospores and finally filled the tip of germination tube (Figs 3b-d). When septum was formed between the germination tube and translucent colloidal substance at its original place, the cells in the germination tube began to divide (Figs 2d-g). Subsequently, they entered the discoid crust stage of early development, with the largest diameter reaching about 310 μm (Figs 2h-i). The adjacent discoid crusts fused with each other and formed larger discoid crusts with 1–5 protrusions in the center of the discoid crusts, then the protrusions grew into upright thalli (Figs 2j-l). After about another 30 d, the upright thalli differentiated into upright branches (Fig. 2m). Upright branches continued to grow for about 50 d and formed young sporelings. Afterwards, they continued to develop into bifurcated sporeling (Figs 2n-o). The development processes of the tetraspores and carpospores were similar.
Figure
3.
The germination and transform progress of protoplast. The colored section was the merge effects of spontaneous fluorescence of chlorophyll and lycopene. a. Carpospores without germination, b. germination tube appeared and the protoplast started to move, c. protoplast continued moving, and d. protoplast filled the tip of germination tube.
The life history of G. constricata included sexual reproduction and asexual propagation (Fig. 4). In sexual reproduction: Gametophytes were dioecious. Spherical and colorless spermatangia were generated from epidermal cells of mature male gametophyte of G. constricata, from which mature spermatia were released. Carpogonial branch ampullar and auxiliary cell ampullar were differentiated from inner cortex of G. constricata. Carpogonial branch contained two cells, hypogynous cell and carpogonium with a long trichogyne at bottom. The ellipsoid auxiliary cells were characterized as bigger and deeper in color compared with other cells. When mature spermatium attached to trichogyne, the nucleus of spermatium got into the carpogonia of the female gametophyte to form zygotes with the egg cells. The fertilized carpogonium fused with the hypogynous cell to form a fusion cell. Afterward, zygotes in fusion cells enter the auxiliary cells to continue developing through the connecting filaments, which was previously formed between fusion cells and auxiliary cells. Then, gonimoblasts were derived from auxiliary cells and continued to divide. Finally, cystocarps emerged. Mature cystocarps scattered over both surfaces of blades, presenting as deep-red spots. When carposporangium matured, carpospores were released through a pore in the outer surface and began to geminate and divide, further formed discoid crusts and turned into upright thalli, finally tetrasporophytes.
Figure
4.
The life history of G. constricata Li et Ding.
In asexual propagation, the inner cortical cells of the sporophyte in G. constricata produced tetrasporocytes, which then formed tetrasporangia. After cruciform division, tetraspores were formed and released. The development process was similar to the carpospores of G. constricata, which finally germinated into female and male gametophytes.
Thus, the life history of G. constricata included homotypic gametophytes (haploid), carposporophytes (diploid), and tetrasporophytes (diploid). The gametophytes were morphologically similar to tetrasporophytes, which indicated that G. constricata belonged to isomorphic alternation of generations.
3.3
Effects of ecological factors on early development
Temperature, irradiance and photoperiod are typical abiotic factors to affect the development and growth of intertidal algae (Fortes and Lüning, 1980; Liu and Dong, 2001; Ouyang et al., 2010; Wei et al., 2013). In order to explore the interaction of ecological factors on the early development of G. constricata, we designed two types of double-factorial experiments include temperature vs. irradiance and temperature vs. photoperiod. The RGR of discoid crust and upright thalli were tested to evaluate growth state.
For the temperature vs. irradiance experiment, we discovered that the RGR of discoid crusts and sporelings increased from 5°C to 20°C and declined from 20°C to 30°C (Fig. 5). The highest RGR of discoid crusts and sporelings occurred at temperature of 20°C and irradiance of 80 μmol photons/(m2·s) (10.583%±0.032% for discoid crusts, 8.869%±0.119% for sporelings). The diameters of discoid crusts and sporelings respectively reached (220±1.414) μm and (3.22±0.117) mm at the optimum condition. The results of double factorial analysis showed that both temperature and irradiance had significant effects on the growth of discoid crusts and sporelings as the single factors. Intriguingly, we found that there was an obvious interaction between temperature and irradiance (Table 2).
Figure
5.
Effects of different temperatures and irradiances on the development of G. constricata Li et Ding discoid crusts (a) and sporelings (b).
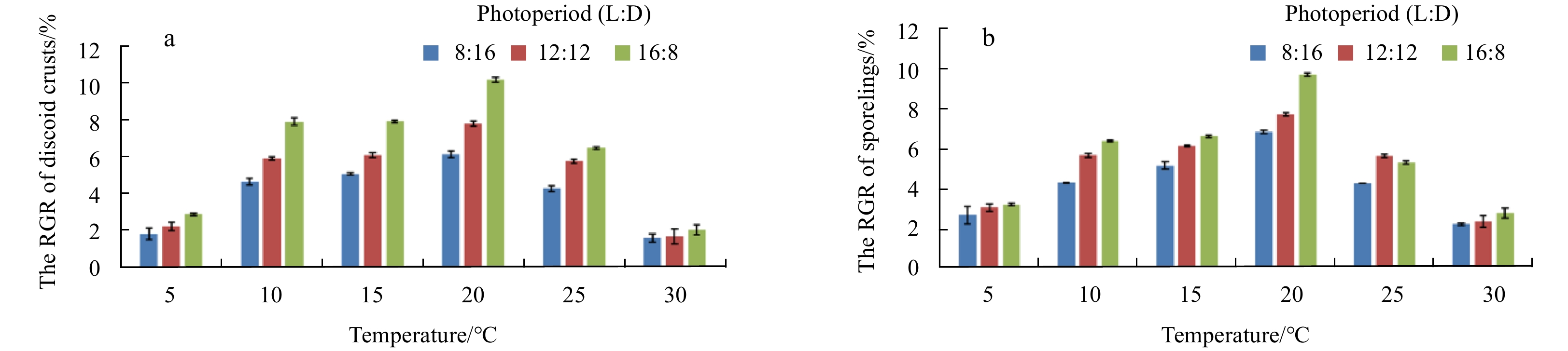
For the temperature vs. photoperiod experiment, the highest RGR of discoid crusts and sporelings were at 20°C in each level of photoperiod. When the photoperiod was 16L:8D, the RGR of discoid crusts and sporelings in each level of temperature reach the maximum except the sporelings cultured at 25°C (Fig. 6). Synthetically, the optimum combination of temperature and photoperiod condition for the growth of discord crusts and sporelings was at 20°C and 16L:8D with the highest RGR (10.251%±0.129% for discoid crusts, 9.670%±0.074% for sporelings). The diameters of discoid crusts and sporelings reached (210±5.437) μm and (3.81±0.085) mm at the optimum condition, respectively. The results of double factorial analysis were similar to that of temperature vs. irradiance, which showed a strong interaction between temperature and photoperiod (Table 3).
Figure
6.
Effects of different temperatures and photoperiods on the development of G. constricata Li et Ding discoid crusts (a) and sporelings (b).
The knowledge of sporogenesis and morphological characteristics including size and type are indispensable to distinguish and identify different species of algae (Shunpei and Hassei, 1947). Here, we observed and recorded the growth and development of spores of G. constricata in detail. The results showed that the type of carpospore development was “mediate discal type” and the life history of G. constricata was the typical isomorphic alternation of generations, which were consistent with G. filicina, type species of Grateloupia, and other Grateloupia species such as G. ramosissima, G. livida, G. tenuis, G. dalianensis, G. huanghaiensis and G. lanceolata (Figs 2 and 4) (Qian, 2014; Zhao, 2012; Song, 2013; Cao et al., 2015; Tian et al., 2017). Meanwhile, we found fusion of discoid crusts, a phenomenon that two or more discoid crusts fused to form a larger discoid crust (Figs 2j-k). As a universal event, fusion of discoid crusts occurred during early development period in many species of Grateloupia and other Rhodophyta (Chen and Ren, 1985; Vera et al., 2008; Li et al., 2010; Wang et al., 2012; Song, 2013; Jiang et al., 2016; Tian et al., 2017). The fusion of multiply discoid crusts could enhance the ability to fix and resist the sour of sea water.
Temperature, photoperiod and Irradiance have been widely reported to affect the growth and development of marine algae and macroalgae (Fortes and Lüning, 1980; Liu and Dong, 2001; Ouyang et al., 2010; Wei et al., 2013). To analyze the ecological habits of G. constricata, we designed two type of double-factorial experiments. We founded that the optimum combination of conditions for the early development of G. constricata spores was as follows: temperature 20°C, irradiance of 80 μmol photons/(m2·s) and photoperiod 16L:8D, which was identical with the results of G. asiatica published by Adharini and Kim (2014). Grateloupia constricata can grow well in the temperature range of 10–25°C, and the relative growth rate of the discoid crusts and sporelings is the largest at 20°C (Fig. 5). The discoid crusts of G. asiatica can grow well at 10–20°C, best at 20°C (Adharini and Kim, 2014). In the suitable temperature range, the spores can develop into discoid crusts quickly and then develop into sporelings. Therefore, the temperature tolerance of the G. constricata is wide enough to be the eurythermal species. When the temperature was under 10°C or above 25°C, the RGR of discoid crusts and sporelings decreased significantly (Fig. 5).
Generally, algae at spore stage are more sensitive to irradiance. High irradiance levels can damage photosynthetic pigments in cells and thereby negatively impact growth (Ouyang et al., 2010). In our study, the optimum irradiance of discoid crusts and sporelings of G. constricata was 80 μmol photons/(m2·s). When the irradiance was 50 μmol photons/(m2·s) and 110 μmol photons/(m2·s), the growth of discoid crusts and sporelings was slower compared with that under the irradiance of 80 μmol photons/(m2·s) (Fig. 5). The reason might be explained as the irradiance of 110 μmol photons/(m2·s) and 50 μmol photons/(m2·s) exceeded and hardly reached the light saturation point of the G. constricata, respectively. Both situations might inhibit the synthesis of chlorophyll a and decrease the photosynthetic rate. Therefore, the energy from photosynthesis was insufficient to maintain algae growth and development. On the other hand, photoperiod adjusted algae growth by changing the illumination time to affect the energy absorbed by the algae growth. As a single ecological factor, longer illumination time might improve the growth rate of G. constricata (Jiang et al., 2009). Our results further verified this and we discovered that the photoperiod of 16L:8D was the optimum condition for the early growth of G. constricata (Fig. 6).
The double factorial analysis showed that the interactions of temperature vs. irradiance and temperature vs. photoperiod both had significant effects on discoid crust development and sporeling growth (P<0.01, Tables 2 and 3). Studies have shown that at moderate temperatures, low irradiance culture condition is more conducive to the development of the discoid crusts of G. yinggehaiensis and G. dalianensis (Zhao, 2012; Wang et al., 2014). However, high irradiance is more suitable for the development of discoid crusts in G. tenuis (Cao et al., 2015). According to the results, we suggested that the interactions of different ecological factors should be considered for cultivation and utilization of G. constricata.
5.
Conclusions
This research reported the life history, especially the early development, and ecological habits of G. constricata. The developmental type of carpospore and tetraspore was mediate discal type. The life history included haploid of gametophyte, diploid of carposporophyte and tetrasporophyte, which showed a typical isomorphic generation alternates because of the similar morphology characteristics of gametophyte and tetrasporophyte. The double factorial analysis of temperature vs. photoperiod and temperature vs. irradiance showed significant interactions on the discoid crusts and sporelings development. The optimum conditions for early growth of G. constricata were temperature 20°C, irradiance 80 μmol photons/(m2·s) and photoperiod 16L:8D.
Adharini R I, Kim H G. 2014. Developmental pattern of crust into upright thalli of Grateloupia asiatica (Halymeniaceae, Rhodophyta). Journal of Applied Phycology, 26(4): 1911–1918. doi: 10.1007/s10811-013-0218-x
[2]
Cao Cuicui, Zhao Fengqin, Guo Shaoru, et al. 2015. Effect of temperature and light on growth of Grateloupia tenuis. Oceanologia et Limnologia Sinica (in Chinese), 46(2): 298–304
[3]
Chen Meiqin, Ren Guozhong. 1985. The development process of sporelings of Gracilaria verrucosa (hudson) papenfuss. Oceanologia et Limnologia Sinica (in Chinese), 16(3): 181–187
[4]
Fang Yuchun, Zhao Xia, Wang Shunchun. 2011. Advances in studies on medicinal alga of Grateloupia C. Agardh. Chinese Journal of Marine Drugs (in Chinese), 30(2): 58–61
[5]
Fortes M D, Lüning K. 1980. Growth rates of North Sea macroalgae in relation to temperature, irradiance and photoperiod. Helgoländer Meeresuntersuchungen, 34(1): 15–29. doi: 10.1007/BF01983538
[6]
Jiang Hongbo, Tian Xiangli, Dong Shuanglin, et al. 2009. Effects of temperature and light intensity on the growth and biochemical composition of Sargassum thunbergii. Chinese Journal of Applied Ecology (in Chinese), 20(1): 185–189
[7]
Jiang Peng, Zhao Shunyu, Li Fang, et al. 2016. Early development of carpospores and life history of Dumontia simplex Cotton. Oceanologia et Limnologia Sinica (in Chinese), 47(1): 151–157
[8]
Li Xiao, Zhao Peng, Wang Gaoge, et al. 2010. Effects of temperature and irradiance on early development of Chondrus ocellatus Holm (Gigartinaceae, Rhodophyta). Chinese Journal of Oceanology and Limnology, 28(3): 508–513. doi: 10.1007/s00343-010-9043-0
[9]
Liu Jingwen, Dong Shuanglin. 2001. Interactions between light and temperature on the growth and levels of chemical constituents of Gracilaria tenuistipitata var. Liui. Journal of Ocean University of Qingdao, 31(3): 332–338
[10]
Ouyang Zhengrong, Wen Xiaobin, Geng Yahong, et al. 2010. The effects of light intensities, temperatures, pH and salinities on photosynthesis of Chlorella. Journal of Wuhan Botanical Research (in Chinese), 28(1): 49–55
[11]
Provasoli L. 1968. Media and prospects for the cultivation of marine algae. In: Watanabe A, Hattori A, eds. Cultures and Collections of Algae. Proceedings of the U.S.–Japan Conference. Hakone, Japan: Japanese Society of Plant Physiology, 63–75
[12]
Qian Shunben. 2014. Phycology (in Chinese). Qingdao: China Ocean University Press, 213–218
[13]
Shunpei I, Hassei K N. 1947. Development of Algae. Tokyo: Hokuryukan, 95–223
[14]
Song Zhimin. 2013. Study on early development of corpospores of Grateloupia livida (in Chinese) [dissertation]. Shantou: Shantou University
[15]
Tian Yilin, Liu Yuwei, Wang Hongwei. 2017. Early development and life cycle of Grateloupia lanceolata. Oceanologia et Limnologia Sinica (in Chinese), 48(1): 113–121
[16]
Vera C, Lobos P, Romo H. 2008. Gametophyte-sporophyte coalescence in populations of the intertidal carrageenophyte Mazzaella laminarioides (Rhodophyta). Journal of Applied Phycology, 20(5): 883–887. doi: 10.1007/s10811-007-9238-8
[17]
Wang Hongwei, Guo Shaoru, Zhang Xiaoming, et al. 2014. Re-identifying Grateloupia yangjiangensis (Rhodophyta, Halymeniaceae) based on morphological observations, life history and rbcL sequence analyses. Acta Oceanologica Sinica, 33(4): 77–84. doi: 10.1007/s13131-014-0450-5
[18]
Wang Gaoge, Jiang Chunmei, Wang Shasha, et al. 2012. Early development of Grateloupia turuturu (Halymeniaceae, Rhodophyta). Chinese Journal of Oceanology and Limnology, 30(2): 264–268. doi: 10.1007/s00343-012-1071-5
[19]
Wei Xiaojiao, Shuai Limei, Lu Bojun. 2013. Effects of temperature and irradiance on filament development of Grateloupia turuturu (Halymeniaceae, Rhodophyta). Journal of Applied Phycology, 25(6): 1881–1886. doi: 10.1007/s10811-013-0018-3
[20]
Xia Bangmei. 2004. Grateloupia C. Agardh (in Chinese). In: Xia Bangmei, ed. Flora Algarum Marinarum Sinicarum (Tomus II Rhodophyta, No. 3 Gelidiales Cryptonemiales Hildenbrandiales). Beijing: Science Press, 87–90
[21]
Zhao Dan. 2012. The life reasearch of four new species of Grateloupia (Halymeniaceae, Rhodophta), from China (in Chinese) [dissertation]. Dalian: Liaoning Normal University
Jae Woo Jung, Qikun Xing, Ji-Sook Park, et al. Effects of salinity and irradiance on early developmental stages of Grateloupia turuturu (Halymeniaceae, Rhodophyta) tetrasporophytes. Algae, 2023, 38(2): 151. doi:10.4490/algae.2023.38.6.2
Yuanyuan Ding, Yao Bian, Huina Wang, Jing Liu, Jingrui Li, Hongwei Wang. Early development, life history and ecological habits of Grateloupia constricata Li et Ding[J]. Acta Oceanologica Sinica, 2020, 39(10): 155-161. doi: 10.1007/s13131-020-1662-5
Yuanyuan Ding, Yao Bian, Huina Wang, Jing Liu, Jingrui Li, Hongwei Wang. Early development, life history and ecological habits of Grateloupia constricata Li et Ding[J]. Acta Oceanologica Sinica, 2020, 39(10): 155-161. doi: 10.1007/s13131-020-1662-5
Figure 1. The field habitat of Grateloupia constricata Li et Ding. a. The female gametophyte of G. constricata Li et Ding (arrow), and b. the male gametophyte of G. constricata Li et Ding (arrow).
Figure 2. The development of carpospores in G. constricata Li et Ding. a. Distribution of carposporophytes (ch represents cystocarp hole), b–c. carpospores, d. transformation of protoplasts, e–h. division of carpospores, i. formation of discoid crust, i–k. fusion of discoid crusts, l. formation of upright thalli, m. upright branches, and n–o. growth of sporeling.
Figure 3. The germination and transform progress of protoplast. The colored section was the merge effects of spontaneous fluorescence of chlorophyll and lycopene. a. Carpospores without germination, b. germination tube appeared and the protoplast started to move, c. protoplast continued moving, and d. protoplast filled the tip of germination tube.
Figure 4. The life history of G. constricata Li et Ding.
Figure 5. Effects of different temperatures and irradiances on the development of G. constricata Li et Ding discoid crusts (a) and sporelings (b).
Figure 6. Effects of different temperatures and photoperiods on the development of G. constricata Li et Ding discoid crusts (a) and sporelings (b).




 DownLoad:
DownLoad:
 DownLoad:
DownLoad:











 DownLoad:
DownLoad: