Yueyun Wang, Hong Cheng, Chunsheng Wang. A new eyeless species of Nicon (Annelida: Nereididae) from the deep Northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2021, 40(12): 20-26. doi: 10.1007/s13131-021-1886-z
Citation:
Jieying Na, Wanying Chen, Dongsheng Zhang, Ruiyan Zhang, Bo Lu, Chengcheng Shen, Yadong Zhou, Chunsheng Wang. Morphological description and population structure of an ophiuroid species from cobalt-rich crust seamounts in the Northwest Pacific: Implications for marine protection under deep-sea mining[J]. Acta Oceanologica Sinica, 2021, 40(12): 79-89. doi: 10.1007/s13131-020-1666-1
Yueyun Wang, Hong Cheng, Chunsheng Wang. A new eyeless species of Nicon (Annelida: Nereididae) from the deep Northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2021, 40(12): 20-26. doi: 10.1007/s13131-021-1886-z
Citation:
Jieying Na, Wanying Chen, Dongsheng Zhang, Ruiyan Zhang, Bo Lu, Chengcheng Shen, Yadong Zhou, Chunsheng Wang. Morphological description and population structure of an ophiuroid species from cobalt-rich crust seamounts in the Northwest Pacific: Implications for marine protection under deep-sea mining[J]. Acta Oceanologica Sinica, 2021, 40(12): 79-89. doi: 10.1007/s13131-020-1666-1
Morphological description and population structure of an ophiuroid species from cobalt-rich crust seamounts in the Northwest Pacific: Implications for marine protection under deep-sea mining
Key Laboratory of Marine Ecosystem Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou 310012, China
2.
Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai), Zhuhai 519080, China
3.
School of Marine Science, China University of Geosciences (Beijing), Beijing 100083, China
4.
School of Oceanography, Shanghai Jiao Tong University, Shanghai 200230, China
5.
State Key Laboratory of Satellite Ocean Environment Dynamics, Hangzhou 310012, China
Funds:
The National Natural Science Foundation of China under contract No. 42076135; the Foundation of China Ocean Mineral Resources R & D Association under contract Nos DY135-E2-2-06 and DY135-E2-2-03; the Project of State Key Laboratory of Satellite Ocean Environment Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, under contract No. SOEDZZ2002; the Scientific Research Fund of the Second Institute of Oceanography, Ministry of Natural Resources, under contract No. JG1528.
Many seamounts are covered with cobalt-rich ferromanganese crusts, and are increasingly attracting interest for the potential extraction of valuable mineral resources from deep seabed environments. However, the impacts of potential mining activities on the vulnerable seamount ecosystem remain unclear. To enhance the understanding of connectivity in benthic invertebrate populations in the Northwest Pacific, several seamounts were surveyed and biological samples collected. In the present study, the ophiuroid species Ophioplinthaca defensor is reported for the first time from four deep seamounts in the Northwest Pacific, and described, providing more detailed morphological diagnosis characters. To assess the population structure of the species between and within seamounts, two mitochondrial markers (COI and 16S) were sequenced. In total, 20 haplotypes from 32 COI sequences and 8 haplotypes from 37 16S sequences were recovered. The star-shaped TCS networks and non-significant pairwise population differences reveal the absence of distinct population structures between and within seamounts. In addition, the O. defensor population seemed to have undergone a demographic expansion in history. This is the first study on the genetic population structure of a benthic invertebrate from seamounts in the Northwest Pacific, and this results suggest a potentially high, long distance dispersal capacity in O. defensor between seamounts, which could inform the development of the Regional Environmental Management Plans for the cobalt-rich crust seamounts in the area.
Nereidids are among the most diverse annelid families, with more than 535 species classified into 43 genera (Bonyadi-Naeini et al., 2017). This family is one of the most diverse groups in the deep sea (Paterson et al., 2009). Comparison with shallow-water species, some morphological and genetic changes has been considered to be an evolutionary adaptation to extreme environment (Zhang et al., 2017). According to Gonzalez et al. (2018), the colonization of Aphroditiformia to deep sea and cave environments seems to be related with the loss of eyes. Many nereidid species belonging to subfamily Nereidinae from deep water are reported as being eyeless. Four Neanthes species (Neanthes abyssorum Hartman, 1967, N. kermadeca (Kirkegaard, 1995), N. typhla (Monro, 1930), N. shinkai Shimabukuro et al., 2017) are known as having no eyes (Shimabukuro et al., 2017). These species are found at depths of more than 2 000 m, with the exception of N. typhla. Neanthes abyssorum was described from Bransfield Strait, Antarctic Ocean; N. kermadeca was from Kermadec Trench, South Pacific Ocean; N. typhla was from off South Georgia Island; N. shinkai was from São Paulo Ridge, Southwest Atlantic Ocean. Nereis anoculopsis Fauchald, 1972 and N. anoculis Hartman, 1960 were from California deep water. They also lacked eyes (Fauchald, 1972; Hartman, 1960). Ceratocephale abyssorum (Hartman and Fauchald, 1971) and Nicon abyssalis at depth of more than 4 000 m are blind too. Nicon, one of the least species rich genera of Nereididae, was described by Kinberg (1866) and including nine species. Those species have a wide distribution from intertidal to deep sea. Subfamily Gymnonereidinae, including Ceratocephale Malmgren, 1867, Gymnonereis Horst, 1919, Micronereides Day, 1963 and Tambalagamia Pillai, 1961, erected by Banse (1977) and characterised by bifid ventral cirri and chaetae forming dense bundles in the anterior 10–15 chaetigers. Fitzhugh (1987) presented the first cladistic analysis of Nereididae and expanded Gymnonereidinae to include all taxa with fully biramous parapodia and with either a smooth or papillate proboscis. Therefore Nicon was included within Gymnonereidinae. Santos et al. (2005) analyzed phylogenetic relationship within Nereididae, and restricted Gymnonereidinae to Banse’s proposal.
In the present manuscript, a new species of eyeless Nicon (Nereididae) from the Northwest Pacific is described. The phylogenetic analysis was carried out to explore the relationship of Nicon inside Nereididae based on four molecular marker: mitochondrial 16S ribosomal RNA gene (16S rDNA) and cytochrome c oxidase subunit I (COI) gene; nuclear 18S ribosomal RNA gene (18S rDNA) and 28S ribosomal RNA gene (28S rDNA).
2.
Materials and methods
The specimen was collected by Hailong III ROV (remotely-operated vehicle) during DY56 Cruise (the 56th China Ocean Scientific Research) in October 2019 and preserved in 95% ethanol solution then deposited in the Sample Repository of Second Institute of Oceanography (RSIO), Ministry of Natural Resources, Hangzhou, China. Specimen was examined under a stereomicroscope (Zeiss Discovery V20). To discern jaws and arrangement of the paragnaths, the withdrawn proboscis dissected. Details of the chaetae were observed using Scanning Electron Microscopy (SEM, HITACHI TM-1000).
The total genomic DNA was extracted from its muscle using DNeasy® Blood and Tissue Kit (QIAGEN, CA, USA) according to the manufacturer’s protocol. DNA concentration used a NanoDrop 2000 Spectrophotometer and a Qubit fluorometer. The qualified DNA was stored in sterilized Milli-Q® water at –80°C before sequencing. The genome sequence was obtained by high-throughput sequencing (PE150) on HiSeqTM X-Ten platform (Illumina, CA, USA) at Zhejiang Tianke High Technology Development Co., Ltd. The de novo assembly was achieved by SPAdes genome assembler (Bankevich et al., 2012). The mitochondrial 16S rDNA and COI gene (cox1), and the nuclear 18S rDNA and 28S rDNA sequences were retrieved from the assembled contigs. The start and end position of the target sequences were checked by the reported universal PCR primer pairs (Norlinder et al., 2012). The 16S rDNA, COI gene, 18S rDNA and 28S rDNA sequences are deposited in GenBank with accession Nos MW525220, MW644966, MW525221 and MW525222, respectively.
The terminals consist of 28 nereidid species (Table 1). Vrijenhoekia balaenophila from family Hesionidae was chosen as the outgroup species. The four marker genes were aligned using the ClustalW algorithm with default settings (15/6.66 as gap/gap length penalties) in Geneious prime software (Biomatters Ltd., New Zealand). The alignments of 16S rDNA, COI gene, 18S rDNA and 28S rDNA sequences were performed separately, and then concatenated into one file after manually trimming unalignable regions. The phylogenetic tree was performed with Maximum-Likelihood (ML) analysis via RAxML GUI v.1.5 software (Stamatakis, 2014; Silvestro and Michalak, 2012) and IQ-TREE package (Nguyen et al., 2015, Chernomor et al., 2016). A partitioned ML analysis was performed in RAxML, with 1 000 bootstrap replicates and the GTRGAMMA model (Lanave et al., 1984; Yang, 1994). The auto-selected best substitution model (-m MFP option) was used in IQ-TREE, also with 1 000 bootstrap replicates. The tree file was visualized and edited in Figtree 1.4.2 (http://tree.bio.ed.ac.uk/software/figtree).
Table
1.
Information and GenBank accession No. of marker genes used in the phylogenetic analysis
Subfamily/family
Taxa
16S rDNA, length/bp
COI gene, length/bp
18S rDNA, length/bp
28S rDNA, length/bp
Nereidinae
Alitta succinea
MN823959, 352
MN823952, 1 515
AY210447, 1 891
AY210464, 3 504
Ceratonereis longiceratophora
−
AY583701, 400
AB106251, 1 642
AF185189, 533
Hediste japonica
LC323064, 416
LC323029, 570
−
LC380658, 804
Hediste diadroma
KX499500, 1 196
KX499500, 1 535
LC323646, 1 698
LC380656, 804
Nectoneanthes oxypoda
−
MN256616, 658
KX290701, 1 793
LC168841, 527
Nereis heterocirrata
KC833492, 462
MN256591, 658
KC840697, 790
−
Nereis pelagica
AY340470, 465
HQ024126, 660
AY340438, 1 795
−
Nereis vexillosa
GU362677, 442
MF121661, 658
DQ790083, 1 829
DQ790043, 3 087
Nereis sp.
MF960765, 1 182
MF960765, 1 534
−
−
Paraleonnates uschakovi
KX462988, 1 191
KX462988, 1 533
−
−
Perinereis wilsoni
LC482173, 422
MN256542, 658
KC840691, 779
−
Perinereis aibuhitensis
KF611806, 1 197
KF611806, 1 534
−
−
Perinereis cultrifera
MN812983, 1 016
MN812983, 1 534
−
−
Perinereis nuntia
JX644015, 1 199
JX644015, 1 537
−
−
Perinereis sp.
MN823971, 1 138
MN823962, 1 536
−
−
Platynereis cf. australis
MN830369, 1 196
MN830369, 1 534
−
−
Platynereis bicanaliculata
MN812984, 1170
MN812984, 1 534
−
−
Platynereis dumerilii
AF178678, 1172
AF178678, 1 534
AY894303, 1 805
−
Pseudonereis variegata
MN855213, 613
MN855134, 1308
KC840693, 779
−
Gymnonereidinae
Ceratocephale abyssorum
GQ426618, 493
GQ426683, 421
GQ426585, 1 709
−
Gymnonereis sp.
KY704332, 472
KY805814, 633
−
−
Laeonereis culveri
KU992689, 1 007
KU992689, 1 536
−
−
Tylorrhynchus heterochaetus
KM111507, 1 236
KM111507, 1 534
−
−
Nicon ablepsiasp. nov.
MW525220,513
MW644966,1 534
MW525221,1 820
MW525222,1 059
Namanereidinae
Namalycastis abiuma
KU351089, 1 019
KU351089, 1 534
−
−
Namalycastis hawaiiensis
LC213728, 521
MN125542, 658
LC213729, 1 781
LC213727, 816
Namalycastis indica
MF959005, 482
MF958995, 660
−
MF959019, 766
Namalycastis jaya
JX483870, 462
JN790067, 699
JX483866, 1 744
−
Hesionidae
Vrijenhoekia balaenophila
JN571884, 512
JN571831, 658
JN571895, 1 777
JN571904, 781
Note: − means the sequence was not reported. The text in bold indicates the measurement results in this paper.
Material examined: Holotype, catalog number B6416500072, KIOST seamount (according to MarineRegions.org, https://www.marineregions.org) in the Northwest Pacific Ocean (Fig. 1), 13.391 5°N, 149.884 7°E, 2 763 m, 5 October 2019.
Figure
1.
Sampling site, KIOST seamount where the sample was collected.
Description: The holotype specimen incomplete, 39 chaetigers, 17.0 mm in length, 1.5 mm wide at chaetiger 1 excluding parapodia (Fig. 2a). Prostomium slightly wider than long with one pair of frontal antennae and one pair of palps (Figs 2b–d). Antennae digitate, same length as palps. Palpostyles sub-conical (Fig. 2d). Eyes absent. Peristomium isometric to chaetiger 1, with four pairs of tentacular cirri; postero-dorsal pair the longest, extending back to posterior margin of 6th chaetiger. Jaws brown, dentate cutting edge with five teeth (Fig. 2e). Oral and maxillary rings without paragnaths and papillae.
Figure
2.Nicon ablepsia sp. nov.: a. dorsal view of the holotype; b. lateral view of the holotype, white triangles mark tentacular cirri; c. dorsal view of head; d. ventral view of head; e. jaws; f. chaetigers 8–17, dorsal view; g. chaetiger 7, end view; and h. chaetiger 7, frontal view. Scale bars: 2.0 mm (a), 0.5 mm (e), 0.05 mm (b, d, f, h).
First two chaetigers only with neuropodia. Biramous chaetigers present from chaetiger 3. Notopodia with dorsal cirri twice as long as notopodial ligule; dorsal cirri inserted in middle of ligule (Figs 2f–h; Fig. 3a). Notopodial ligule and notopodial lobe sub-conical; ligule slightly shorter than lobe. Neuropodial ligule conical, stouter than lobe. Ventral cirri inserted at base of ligule, slightly shorter than it. Notochaetae present from chaetiger 3, homogomph spinigers (Fig. 3d). Homogomph spinigers and heterogomph falcigers present in neurochaetae dorsal and ventral fascicle (Figs 3e and f). Acicular chaetae black in color. Notoaciculae visible on epidermis of notopodial lobe with curved distal ends on middorsal side of notopodial lobe (Figs 2f and 3b). Neoroaciculae straight with curved tips (Fig. 3e).
Figure
3.
Parapodia and chaetae (a−d, SEM; e−g, optical picture). a. Parapodium; b. notopodial lobe, arrow shows curved tip of notoacicula; c. neurochaetae, homogomph spinigers and heterogomph falcigers; d. notochaetae, homogomph spinigers; e. neuropodium with neurochaetae; f. heterogomph falcigers; and g. notochaetae, homogomph spinigers. Scale bars: 0.5 mm (a), 0.1 mm (b−d).
Total segmental number and pygidium unknown as the specimen incomplete.
Etymology: The specific name ablepsia come from Latin and refers to the eyeless character of this new species.
Distribution: Known only for type locality.
Molecular analysis: The length of 16S rDNA, COI gene, 18S rDNA and 28S rDNA sequences were confirmed by respective sequence of PCR primers (Norlinder et al., 2012). The primer-sequence alignments of PCR primers located in relevant position of marker genes correctly. The partial 16S rDNA, the complete COI gene, the partial 18S rDNA and 28S rDNA sequences of N. ablepsia sp. nov. (voucher B6416500072) contained 1 820 bp, 1 059 bp, 513 bp and 1 534 bp length, respectively.
Genera Namalycastis and Tylorrhynchus formed a well-supported clade, which was sister to the group of Nereidinae + Nicon + Laeonereis clade. Neither the Gymnonereidinae sensu Fitzhugh (1987) nor Nereidinae were recovered as monophyleticis as Nicon and Laeonereis being clustered with genera from Nereidinae. The Nicon ablepsia sp. nov. and Alitta succinea showed different topological cluster in the RaxML tree (Fig. 4) and IQ-tree (Fig. A1 in Appendix) with very low support value.
Figure
4.
The Maximum-Likelihood phylogenetic tree of 29 species based on the concatenated dataset of 16S rDNA, COI gene, 18S rDNA and 28S rDNA sequences with 1 000 bootstrap replicates. Bootstrap support values (calculated by RAxML software) are indicated adjacent to each node. The red solid dot shows the inconsistent topology cluster between the tree constructed by RAxML and IQ-TREE. All the other nodes were identical.
The genus Nicon is characterized by having all notochaetae homogomph spinigers and neurochaetae falcigers, lacking paragnaths or papillae on oral and maxillary ring. de León-González and Trovant (2013) recognized eleven Nicon species. Later, N. polaris was transferred to Kainonereis since it bears elytriform structures (Conde-Vela et al., 2018). Nicon sinica Wu & Sun, 1979 was transferred to Sinonereis because it is a junior synonym of Sinonereis heteropoda based on affinities in chaetal and parapodial features (Conde-Vela and Wu, 2019). The new species lacks eyes, therefore, is easily distinguished from other species of Nicon. Nicon ablepsia sp. nov. is similar to N. abyssalis by absence of eyes but differs from the latter in the position of the antennae, length of tentacular cirri and relative length between peristomium and chaetiger 1. The antennae of N. abyssalis are inserted closely together in the frontal margin of the prostomium, and the bases of the antennae are in contact (Hartman, 1967). The antennae of N. ablepsia sp. nov. are separate from each other. Tentacular cirri are short in N. abyssalis (reaching chaetiger 2), but long in N. ablepsia sp. nov., reaching chaetiger 6. The peristomium of N. abyssalis is prolonged and nearly twice as long as chaetiger 1, while equal to the length of chaetiger 1 in N. ablepsia sp. nov.
The phylogeny of the Nereididae and the higher taxonomic relationships within it has been explored through molecular approaches recently (Alves et al., 2020). However, the molecular phylogenetic relationship between Nicon and other Nereidid genera has not been investigated yet. Santos et al. (2005) assessed relationships of nereidids in a morphology-based parsimony analysis of 41 terminal taxa. Gymnonereidinae was restricted to Ceratocephale, Gymnonereis, Tambalagamia and Micronereides. Systematic status of Nicon was not resolved in Santos’s results. In our tree, Niconablepsia sp. nov. is closely to Alitta succinea with low support. Alitta is characterized by conical paragnaths on maxillary and oral ring, and dorsal cirrus mid-dorsally to subterminally attached to dorsal notopodial ligule on posterior chaetigers (Bakken and Wilson, 2005). Niconablepsia sp. nov. has no paragnaths and dorsal cirrus attached to base of notopodium.
The explicit systematic status of Nicon is still undetermined due to the type species of Nicon, and some of its closely related genera are not included in the present analysis. Therefore, more extensive taxon coverage are required in further studies to elucidate the systematic status of Nicon.
The sequence data reported can provide base information for molecular phylogenetic research of Family Nereididae in the future, and can also supply the basic information for designing primers. A taxonomic key to species of Nicon is provided here.
– Heterogomph falcigers absent, with sesquigomph falcigers in infracicular position, dorsal ligule long and thin on median and posterior parapodia........................................ N. orensanzi
4. With homogomph falcigers in neuropodial subacicular posi tion............................................................................... N. rotunda
– Homogomph falcigers lacking.............................. N. japonica
6. Dorsal ligule cirriform, reduced in posterior chaetigers; falci gers with prolonged blade................................... N. abysssalis
– Dorsasl ligule subtriangular, similar in size throughout; falci gers with long, anteriorly blunt blade distinctly serrated along inner margin............................................................. N. yaquinae
7. Eyes present; tentacular cirri reaching chaetiger 5; supra and infracicular sesquigomph falcigers present......... N. pettibonae
– Eyes absent; tentacular cirri reaching chaetiger 6; supra and infracicular sesquigomph falcigers absent.. N. ablepsia sp. nov.
8. Longest pair of tentacular cirri partially annulated on distal end; falcigers with long blade, denticulate along inner margin .................................................................................... N. maculate
– All tentacular cirri annulated, with cylindrical articles; falci gers with short blades, denticles on proximal inner margin ............................................................................. N. moniloceras
Acknowlegements
We thank the crew of R/V Dayang Yihao and technical staff of Hailong III ROV for their generous help during the investigation.
A1.
The Maximum-Likelihood tree constructed by IQ-TREE. Numbers near the branch nodes refer to SH-aLRT/ UFBoot support values (all based on 1 000 replicates). The red solid dot showed the inconsistent topology cluster between the tree constructed by RAxML and IQ-TREE. The partitioned scheme and best model information was auto selected by IQ-TREE as follows: Part 1 (1–1 158 nt, 16S rDNA), GTR+F+I+G4 model; Part 2 (1 159–2 697 nt, COI gene), TIM2+F+R4 model; Part 3 (2 698–4 600 nt, 18S rDNA), TIM2e+FQ+I+G4 model; and Part 4 (4 601–5 559 nt, 28S rDNA), TN+F model.
Baco A R, Etter R J, Ribeiro P A, et al. 2016. A synthesis of genetic connectivity in deep-sea fauna and implications for marine reserve design. Molecular Ecology, 25(14): 3276–3298. doi: 10.1111/mec.13689
[2]
Balbar A C, Metaxas A. 2019. The current application of ecological connectivity in the design of marine protected areas. Global Ecology and Conservation, 17: e00569. doi: 10.1016/j.gecco.2019.e00569
[3]
Boissin E, Stöhr S, Chenuil A. 2011. Did vicariance and adaptation drive cryptic speciation and evolution of brooding in Ophioderma longicauda (Echinodermata: Ophiuroidea), a common Atlanto-Mediterranean ophiuroid?. Molecular Ecology, 20(22): 4734–4755
[4]
Boschen R E, Tyler P A, Copley J T. 2013. Distribution, population structure, reproduction and diet of Ophiolimna antarctica (Lyman, 1879) from Kemp Caldera in the Southern Ocean. Deep-Sea Research Part II: Topical Studies in Oceanography, 92: 27–35. doi: 10.1016/j.dsr2.2013.02.005
[5]
Castelin M, Lorion J, Brisset J, et al. 2012. Speciation patterns in gastropods with long-lived larvae from deep-sea seamounts. Molecular Ecology, 21(19): 4828–4853. doi: 10.1111/j.1365-294X.2012.05743.x
[6]
Cho W, Shank T M. 2010. Incongruent patterns of genetic connectivity among four ophiuroid species with differing coral host specificity on North Atlantic seamounts. Marine Ecology, 31(S1): 121–143
[7]
Clark M R, Bowden D A, Rowden A A, et al. 2019. Little evidence of benthic community resilience to bottom trawling on seamounts after 15 years. Frontiers in Marine Science, 6: 63. doi: 10.3389/fmars.2019.00063
[8]
Clark M R, Rowden R A, Schlacher T, et al. 2010. The ecology of seamounts: structure, function, and human impacts. Annual Review of Marine Science, 2: 253–278. doi: 10.1146/annurev-marine-120308-081109
[9]
Edgar R C. 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res, 32(5): 1792–1797, doi: 10.1093/nar/gkh340
[10]
Etter R J, Boyle E E, Glazier A, et al. 2011. Phylogeography of a pan-Atlantic abyssal protobranch bivalve: Implications for evolution in the Deep Atlantic. Molecular Ecology, 20(4): 829–843. doi: 10.1111/j.1365-294X.2010.04978.x
[11]
Excoffier L, Lischer H E L. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Molecular Ecology Resources, 10(3): 564–567. doi: 10.1111/j.1755-0998.2010.02847.x
[12]
Fu Yunxin. 1997. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics, 147(2): 915–925. doi: 10.1093/genetics/147.2.915
[13]
Galaska M P, Sands C J, Santos S R, et al. 2016. Geographic structure in the Southern Ocean circumpolar brittle star Ophionotus victoriae (Ophiuridae) revealed from mtDNA and single- nucleotide polymorphism data. Ecology and Evolution, 7(2): 475–485
[14]
Galaska M P, Sands C J, Santos S R, et al. 2017. Crossing the Divide: Admixture Across the Antarctic Polar Front Revealed by the Brittle Star Astrotoma agassizii. The Biological Bulletin, 232(3): 198–211. doi: 10.1086/693460
[15]
Harpending H C. 1994. Signature of ancient population growth in a low-resolution mitochondrial DNA mismatch distribution. Human Biology, 66(4): 591–600
[16]
Havermans C, Sonet G, D’Acoz C D, et al. 2013. Genetic and morphological divergences in the cosmopolitan deep-sea amphipod Eurythenes gryllus reveal a diverse abyss and a bipolar species. PLoS ONE, 8(9): e74218. doi: 10.1371/journal.pone.0074218
[17]
Hein J R, Bohrson W A, Schulz M S, et al. 1992. Variations in the fine-scale composition of a central pacific ferromanganese crust: paleoceanographic implications. Paleoceanography and Paleoclimatology, 7(1): 63–77
[18]
Hilário A, Metaxas A, Gaudron S M, et al. 2015. Estimating dispersal distance in the deep sea: challenges and applications to marine reserves. Frontiers in Marine Science, 2: 6. doi: 10.3389/fmars.2015.00006
[19]
Hunter R L, Halanych K M. 2008. Evaluating connectivity in the brooding brittle star Astrotoma agassizii across the Drake Passage in the Southern Ocean. Journal of Heredity, 99(2): 137–148. doi: 10.1093/jhered/esm119
[20]
Hunter R L, Halanych K M. 2010. Phylogeography of the Antarctic planktotrophic brittle star Ophionotus victoriae reveals genetic structure inconsistent with early life history. Marine Biology, 157(8): 1693–1704. doi: 10.1007/s00227-010-1443-3
[21]
ISA. 2018. Towards the development of a regional environmental management plan for cobalt-rich ferromanganese crusts in the Northwest Pacific Ocean. ISA Technical Study No. 23. Kingston, Jamaica: International Seabed Authority
[22]
ISBA/19/LTC/8. 2013. Recommendations for the guidance of contractors for the assessment of the possible environmental impacts arising from exploration for marine minerals in the area. Jamaica: International Seabed Authority
[23]
Jennings R M, Etter R J, Lynn F, et al. 2013. Population differentiation and species formation in the deep sea: the potential role of environmental gradients and depth. PloS ONE, 8(10): e77594. doi: 10.1371/journal.pone.0077594
[24]
Kaiho K, Hasegawa T. 1994. End-Cenomanian benthic foraminiferal extinctions and oceanic dysoxic events in the northwestern Pacific Ocean. Palaeogeography, Palaeoclimatology, Palaeoecology, 111(1–2): 29–43. doi: 10.1016/0031-0182(94)90346-8
[25]
Koehler R. 1930. Ophiures recueillies par le Docteur Th. Mortensen dans les Mers d’Australie et dans I’Archipel Malais. Papers from Dr. Th. Mortensen’s Pacific Expedition 1914-16. LIV. Videnskabelige Meddelelser fra Dansk naturhistorisk Forening, 89: 1–295
[26]
Leigh J W, Bryant D. 2015. Popart: full-feature software for haplotype network construction. Methods in Ecology and Evolution, 6(9): 1110–1116. doi: 10.1111/2041-210X.12410
[27]
Manel S, Loiseau N, Andrello M, et al. 2019. Long-distance benefits of marine reserves: myth or reality?. Trends in Ecology & Evolution, 34(4): 342–354
[28]
Martino S, Parson L M. 2012. A comparison between manganese nodules and cobalt crust economics in a scenario of mutual exclusivity. Marine Policy, 36(3): 790–800. doi: 10.1016/j.marpol.2011.11.008
[29]
Miller K J, Gunasekera R M. 2017. A comparison of genetic connectivity in two deep sea corals to examine whether seamounts are isolated islands or stepping stones for dispersal. Scientific Reports, 7: 46103. doi: 10.1038/srep46103
[30]
Morgan N B, Cairns S, Reiswig H, et al. 2015. Benthic megafaunal community structure of cobalt-rich manganese crusts on Necker Ridge. Deep-Sea Research Part I: Oceanographic Research Papers, 104: 92–105. doi: 10.1016/j.dsr.2015.07.003
[31]
O’Driscoll R L, Clark M R. 2005. Quantifying the relative intensity of fishing on New Zealand seamounts. New Zealand Journal of Marine and Freshwater Research, 39(4): 839–850. doi: 10.1080/00288330.2005.9517356
[32]
O’Hara T D. 2007. Seamounts: centres of endemism or species-richness for ophiuroids?. Global Ecology and Biogeography, 16(6): 720–732. doi: 10.1111/j.1466-8238.2007.00329.x
[33]
O’Hara T D, Consalvey M, Lavrado H P, et al. 2010. Environmental predictors and turnover of biota along a seamount chain. Marine Ecology, 31(S1): 84–94
[34]
O’Hara T D, England P R, Gunasekera R M, et al. 2014. Limited phylogeographic structure for five bathyal ophiuroids at continental scales. Deep-Sea Research Part I: Oceanographic Research Papers, 84: 18–28. doi: 10.1016/j.dsr.2013.09.009
[35]
O’Hara T D, Smith P J, Mills V S, et al. 2013. Biogeographical and phylogeographical relationships of the bathyal ophiuroid fauna of the Macquarie Ridge, Southern Ocean. Polar Biology, 36(3): 321–333. doi: 10.1007/s00300-012-1261-9
[36]
O’Hara T D, Stöhr S. 2006. Deep water Ophiuroidea (Echinodermata) of New Caledonia: Ophiacanthidae and Hemieuryalidae. In: Richer de Forges B, Justine J L, eds. Tropical Deep Sea Benthos (Mémoires du Muséum national d’Histoire naturelle 193). Paris, France: Museum of Natural History of France, 24: 33–141
[37]
Palumbi S R. 1996. Nucleic acidS II: the polymerase chain reaction. In: Hilli D M, Moritz C, Mable B K, eds. Molecular Systematics. 2nd ed. Sunderland, MA, USA: Sinauer Associates, 205–247
[38]
Palumbi S R. 2003. Population genetics, demographic connectivity, and the design of marine reserves. Ecological Applications, 13(S1): 146–158. doi: 10.1890/1051-0761(2003)013[0146:PGDCAT]2.0.CO;2
[39]
Pérez-Portela R, Almada V, Turon X. 2012. Cryptic speciation and genetic structure of widely distributed brittle stars (Ophiuroidea) in Europe. Zoologica Scripta, 42(2): 151–169
[40]
Rozas J, Ferrer-Mata A, Sánchez-DelBarrio J C, et al. 2017. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Molecular Biology and Evolution, 34(12): 3299–3302. doi: 10.1093/molbev/msx248
[41]
Samadi S, Bottan L, Macpherson E, et al. 2006. Seamount endemism questioned by the geographic distribution and population genetic structure of marine invertebrates. Marine Biology, 149(6): 1463–1475. doi: 10.1007/s00227-006-0306-4
[42]
Sands C J, O’Hara T D, Barnes D K A, et al. 2015. Against the flow: evidence of multiple recent invasions of warmer continental shelf waters by a Southern Ocean brittle star. Frontiers in Ecology and Evolution, 3: 63. doi: 10.3389/fevo.2015.00063
[43]
Schlacher T A, Baco A R, Rowden A A, et al. 2014. Seamount benthos in a cobalt-rich crust region of the central Pacific: conservation challenges for future seabed mining. Diversity and Distributions, 20(5): 491–502. doi: 10.1111/ddi.12142
[44]
Shank T M. 2010. Seamounts: deep-ocean laboratories of faunal connectivity, evolution, and endemism. Oceanography, 23(1): 108–122. doi: 10.5670/oceanog.2010.65
[45]
Taboada S, Pérez-Portela R. 2016. Contrasted phylogeographic patterns on mitochondrial DNA of shallow and deep brittle stars across the Atlantic-Mediterranean area. Scientific Reports, 6: 32425. doi: 10.1038/srep32425
[46]
Taboada S, Riesgo A, Wiklund H, et al. 2018. Implications of population connectivity studies for the design of marine protected areas in the deep sea: An example of a demosponge from the Clarion-Clipperton Zone. Molecular Ecology, 27(23): 4657–4679. doi: 10.1111/mec.14888
[47]
Tajima F. 1989. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics, 123(3): 585–595. doi: 10.1093/genetics/123.3.585
[48]
Taylor M L, Roterman C N. 2017. Invertebrate population genetics across Earth’s largest habitat: The deep-sea floor. Molecular Ecology, 26(19): 4872–4896. doi: 10.1111/mec.14237
[49]
Thatje S, Hillenbrand C D, Mackensen A, et al. 2008. Life hung by a thread: endurance of Antarctic fauna in glacial periods. Ecology, 89(3): 682–692. doi: 10.1890/07-0498.1
[50]
Yesson C, Clark M R, Taylor M L, et al. 2011. The global distribution of seamounts based on 30 arc seconds bathymetry data. Deep-Sea Research Part I: Oceanographic Research Papers, 58(4): 442–453. doi: 10.1016/j.dsr.2011.02.004
[51]
Zeng Cong, Clark M R, Rowden A A, et al. 2019. The use of spatially explicit genetic variation data from four deep-sea sponges to inform the protection of Vulnerable Marine Ecosystems. Scientific Reports, 9: 5482. doi: 10.1038/241598-019-41877-9
Tulio F. Villalobos-Guerrero, Sonja Huč, Ekin Tilic, et al. A remarkable new deep-sea nereidid (Annelida: Nereididae) with gills. PLOS ONE, 2024, 19(3): e0297961. doi:10.1371/journal.pone.0297961
2.
Deyuan Yang, Sheng Zeng, Zhi Wang, et al. Molecular systematics of Perinereis and an investigation of the status and relationships of the cultured species Perinereis wilsoni Glasby & Hsieh, 2006 (Annelida, Nereididae). Zoosystematics and Evolution, 2024, 100(4): 1297. doi:10.3897/zse.100.127201
3.
Robin S. Wilson, Christopher J. Glasby, Torkild Bakken. The Nereididae (Annelida) – diagnoses, descriptions, and a key to the genera. ZooKeys, 2023, 1182: 35. doi:10.3897/zookeys.1182.104258
4.
Andrei V. Grischenko. New deep-water Bryozoa from the Magellan Seamounts, tropical western Pacific, with a global review of bryozoans associated with ferromanganese crusts. Progress in Oceanography, 2022, 205: 102827. doi:10.1016/j.pocean.2022.102827
Yueyun Wang, Hong Cheng, Chunsheng Wang. A new eyeless species of Nicon (Annelida: Nereididae) from the deep Northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2021, 40(12): 20-26. doi: 10.1007/s13131-021-1886-z
Yueyun Wang, Hong Cheng, Chunsheng Wang. A new eyeless species of Nicon (Annelida: Nereididae) from the deep Northwest Pacific Ocean[J]. Acta Oceanologica Sinica, 2021, 40(12): 20-26. doi: 10.1007/s13131-021-1886-z
Figure 1. Map of the study seamounts in the Northwest Pacific (a) and sampling sites in the Weijia Guyot (b).
Figure 2. Morphological characters of Ophioplinthaca defensor (RSIO410602). a. Dorsal view; b. ventral view of disc; c. the interradial characters of ventral side; d. dorsal view of arms, proximal part; e. ventral view of arm, proximal part; f. dorsal view of arms, distal part. AD: adoral shield; AS: arm spines; DAP: dorsal arm plate; DS: disc spines; GS: genital slits; J: jaw; OP: oral papillae; OS: oral shield; RS: radial shield; VAP: ventral arm plate. Scale bars: 2 mm.
Figure 3. Scanning electron microscope photographs of arm skeletal characters and disc spines of Ophioplinthaca defensor. a–f. Arm skeletal characters from adult paratype (RSIO410602): proximal view of vertebrae; distal view of vertebrae; lateral view of vertebrae; ventral view of vertebrae; dorsal view of vertebrae; external view of lateral arm plate. g–i. Different types of disc spines, large cylindrical granules from adult paratype RSIO410602; conical or round granules from adult paratype RSIO410617; disc spines cylindrical with flaring thorny apex from juvenile paratype RSIO410502. mo: muscle opening; no: nerve opening. Arrows: d. dorsal side; v. ventral side; dist. distal side; prox. proximal side. Scale bars: 200 μm.
Figure 4. Variation in morphological characters of Ophioplinthaca defensor. a–c. Adult (RSIO410617) from the Weiija Guyot: dorsal view of disc; disc granules; ventral view of disc. d–f. Adult (RSIO358101) from the Caiwei Guyot: dorsal view of disc; disc granules; ventral view of disc. g–i. Juvenile (RSIO410502) from the Weijia Guyot: dorsal view of disc; disc granules; ventral view of disc. Scale bars: 2 mm (a, c, d, f), 500 μm (b, e, g–i).
Figure 5. COI (a) and 16S (b) TCS haplotype networks. Each circle represents a haplotype and its diameter is proportional to the frequency of the haplotype. Median vectors (black circles, which represent missing intermediates haplotypes) are added to the network.
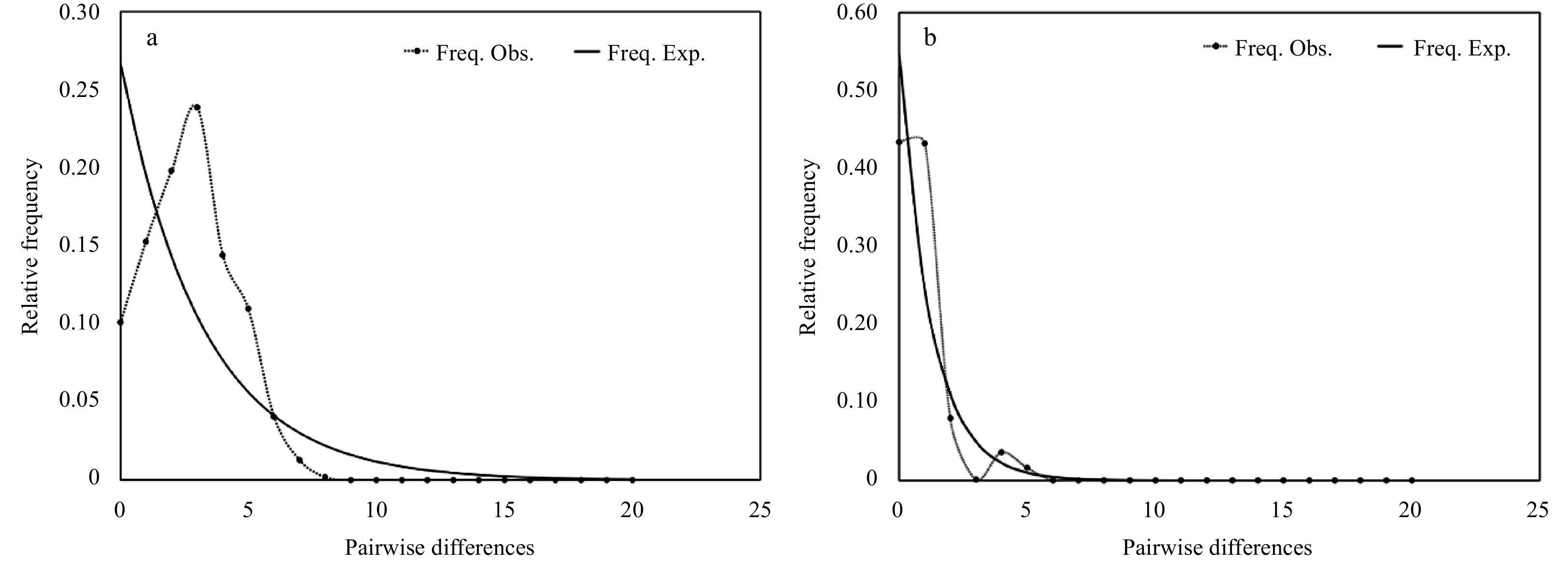
Figure 6. Pairwise mismatch distribution for all COI (a) and 16S (b) sequences of Ophioplinthaca defensor from the Weijia Guyot.
Figure 7. Different sponge species with Ophioplinthaca defensor individuals attached in the Weijia Guyot. a. Farreidae sp. 1; b. Farreidae/Euretidae sp.; c. Corbitellinae sp.; d. Farreidae sp. 2. Red arrows indicate Ophioplinthaca defensor.



 DownLoad:
DownLoad:

 DownLoad:
DownLoad: