| Citation: | Wenlin Wu, Hongyun Li, Tiantian Ma, Xiaobo Zhang. Characterization of DNA polymerase δ from deep-sea hydrothermal vent shrimp Rimicaris exoculata[J]. Acta Oceanologica Sinica, 2021, 40(8): 168-175. doi: 10.1007/s13131-021-1823-1

|
Deep-sea hydrothermal vents (DSHV) are spatially and temporally unstable environment because of the frequent volcanic and tectonic events that often lead to the destruction or birth of these habitats. Therefore, the habitats of DSHV are extreme and unique with the fluids around DSHV enriched in sulphide, heavy metals, high pressure, and low dissolved oxygen (Hutnak et al., 2008; Konn et al., 2015). Biotic communities around the hydrothermal vents have developed strategies, mostly a combination of physiological and behavioral adaptations, to adapt or avoid the extreme conditions of their habitats. Generally the enzymatic activities of the major pathways of energy metabolism in vent animals are qualitatively and quantitatively similar to those of phylogenetically related shallow-living marine species, suggesting that the types of energy metabolism pathways and the potential flux rates through these pathways are similar in both groups (Haki and Rakshit, 2003). Among the vent faunas, there lives a special invertebrate, the vent shrimp Rimicaris exoculata, which belongs to the family Alvinocarididae (Williams and Rona, 1986). Shrimp R. exoculata is the predominant species of most hydrothermal vents along the Mid-Atlantic ocean ridge and has also been discovered in the Mid-Cayman Spreading Center and Southwest Indian Ocean Ridge (Zhang et al., 2017). It forms dense and moving aggregates, located closely along chimney walls in the gradient between hydrothermal fluids and cold oxygenated ambient seawater, in a temperature range between 3°C and 25°C (Komaï and Segonzac, 2008; Ravaux et al., 2009; Le Bloa et al., 2017). To date, the mechanism underlying their environmental adaptation and survival is unclear.
As well known, DNA replication is a basic and conserved event in cells that has a fundamental function for genetics and evolution (Kunkel and Burgers, 2017). Many DNA polymerases and protein factors are involved in DNA replication process with high fidelity. Therefore, DNA polymerases of vent shrimp R. exoculata may have evolved some unique features to fit the extreme environment of deep-sea hydrothermal vent. At present, however, DNA polymerase of deep-sea animals has not been investigated. In eukaryotes, DNA polymerase δ (Polδ) is an essential protein in chromosomal DNA replication and multiple DNA repair processes, depending on its DNA-directed polymerase activity and 3′-5′ exonuclease activity (Swan at al., 2009). During lagging strand DNA replication Polδ performs three essential functions in the process. Firstly, Polδ catalyzes extension of the short Okazaki fragments generated by DNA Polα via DNA-directed DNA synthesis to fill the gap between two successive Okazaki fragments (Maga et al., 2001; Burgers and Kunkel, 2017). Secondly, Polδ checks the mis-incorporated bases and correct them via its 3′-5′ exonuclease activity, thus ensuring a relative high fidelity in copying the template strand (Fortune et al., 2005). Finally, during the maturation of Okazaki fragment, Polδ catalyzes strand displacement DNA synthesis through the downstream Okazaki fragment to allow the production of 5′-flaps as substrates for FEN1 endonucleases (Jin et al., 2001). In addition to its role in DNA replication, Polδ also involves in DNA repair and recombination (Hübscher et al., 2002).
Previous studies indicated that Polδ is composed of three subunits in yeast, including the catalytic subunit Pol3p (corresponding to mammalian p125, POLD1), the structural subunits Pol31p (corresponding to mammalian p50, POLD2) and Pol32p (corresponding to mammalian p68, POLD3) (Johnson et al., 2012). Mammals Polδ consists another smallest subunit p12 (POLD4) (Prindle and Loeb, 2012). POLD1 is a key component of DNA Pol δ catalytic subunit and plays an important role in cell growth and differentiation. Accumulation evidences in the past decade have linked germline and sporadic mutations in POLD1 with human pathologies. Genome sequencing has identified mutations in POLD1 as the cause of some inherited cancers (Rayner et al., 2016). In addition, mutations in POLD1 have been found in the developmental disorders of mandibular hypoplasia, deafness, progeroid features and lipodystrophy and atypical Werner syndrome (Weedon et al., 2013).
The present study describes the cloning and expression of vent shrimp R.exoculata POLD1 and the characterization of its activity. In addition, the in vitro inhibitory effects of some metal and detergents were evaluated on the activity of POLD1.
Vent shrimp R. exoculata was collected by a television grab from a newly discovered deep-sea hydrothermal vent (15°09′S, 13°21′W, 2 800 m in depth) in the South Mid-Atlantic Ridge during legIII of the 26th cruise of “China Ocean”. In our previous study (data not published), the vent shrimp genome was partially sequenced. In the partial genomic sequence of vent shrimp, the Polδ catalytic subunit gene was found. Based on the sequence, the POLD1 gene was cloned using sequence-specific primers (5′-GATCCA GTTATTCAAATTGC-3′ and 5′-CACGGCTGGTGCATATAACTTC-3′). The muscle of a vent shrimp was homogenized in liquid nitrogen. Then total RNAs were extracted with Tissue/Cell RNA Rapid Extraction Kit (Aidlab Biotechnologies, Beijing, China). The first stranded cDNA was synthesized using oligo dT-anchor primer and MMLV reverse transcriptase (TaKaRa, Dalian, China). Subsequently PCR was conducted to clone POLD1 of vent shrimp.
To obtain the full-length cDNA of POLD1, 5′ and 3′ RACEs (rapid amplification of cDNA ends) were performed using 5′ and 3′ RACEs kit according to the manufacturer’s instructions (Roche, Basel, Switzerland). For 3′ RACE, the first round of PCR was carried out with specific primers (5′-GTTCCACTAAATTCAGCATCAG-3′ and 5′-GACCA CGCGTATCGATGTCGACTTTTTTTTTTTTTTTT-3′). The nested PCR was performed using nested primer (5′-GAGTGAGATTTACATGACC-3′) and PCR anchor primer (5′-GACCACGCGTATCGATGTCGAC-3′). For 5′ RACE, the reverse transcription was conducted with a specific primer (5′-CTTTAACTCTTCCAAGAAATGAAAAAG-3′). The first round of PCR was performed using a specific primer (5′-GTCCACATGGCGCAAAAATGTTGC-3′) and the oligo dT-anchor primer. Then the nested PCR was conducted with nested primer (5′-GACTGGACCTTGGACCCC AC-3′) and the PCR-anchor primer.
Phylogenetic analysis was restricted to full-length amino acid sequence of POLD1 that could be aligned unambiguously in all sequences. Eight crustaceans were used for the phylogenetic analysis of POLD1, including R. exoculata, Triops granarius (GenBank accession no BAO20829.1), Oratosquilla oratoria (GenBank accession no BAO20826.1), Hyalella azteca (GenBank accession no XP 018015591.1), Cyclops vicinus (GenBank accession no BAO20825.1), Branchinella kugenumaensis (GenBank accession no BAO20830.1), Procambarus clarkii (GenBank accession no BAO20827.1) and Daphnia pulicaria (GenBank accession no BAO20828.1). The evolution was analyzed using the Neighbor-Joining method. The optimal tree with the sum of branch length (0.944 9) was shown. The tree was drawn to scale with branch lengths (next to the branches) in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were calculated using the Poisson correction method and were in the units of the number of amino acid substitutions per site. All positions containing gaps and missing data were eliminated. There were a total of 841 positions in the final dataset. Evolutionary analyses were conducted in MEGA7 (Kumar et al., 2016).
The Polδ catalytic subunit gene was cloned into pET28a vector and expressed as a His tag fusion protein in Escherichia coli BL21 (Invitrogen, Carlsbad, CA, USA). Protein expression was induced by growing the bacteria to an OD600 of 0.6 and adding isopropyl-β-d-thiogalactoside (IPTG) to a final concentration of 1 mmol/L at 37°C for 4 h. The bacteria were disrupted by sonication in lysis buffer (50 mmol/L sodium phosphate, 500 mmol/L NaCl, 1 mmol/L DL-Dithiothreitol, 0.1 mmol/L phenylmethylsulfonyl fluoride, pH 7.9). The lysate was then centrifuged at 18 000× g for 25 min. The supernatant was incubated with Ni sepharoseTM 6 Fast Flow (GE Healthcare, Pittsburgh, PA, USA) overnight at 4°C. After washes with lysis buffer, the protein was eluted with elution buffer (20 mmol/L Tris-HCl, 200 mmol/L NaCl, 500 mmol/L Imidazole, pH 8.0).
The DNA polymerase activity was measured as described before with some modifications (Tveit and Kristensen, 2001). Briefly, the oligonucleotide template (5′-CTGTTTCG-CTTCAGAAGAGACGGTGACCGAAAACGCATACTCCTCCAACACAATGCGTCCATTGACCGCCA-3′) (6 μmol/L), the oligonucleotide primer (5′-CTCTTCTGAAG CGAAA-3′) (8 μmol/L) and the annealing buffer (10 mmol/L Tris-HCl, 30 mmol/L NaCl, 1 mmol/L EDTA, pH 7.5) at a total volume of 100 μL were heated at 95°C for 10 min, followed by slowly cooling to room temperature. Subsequently the annealed primer-template mixture, 1 μg/mL POLD1 and the reaction buffer (40 mmol/L Tris-HCl, 6 mmol/L MgCl2, 100 μmol/L dNTP, 100 μg/mL bovine serum albumin, 1 mmol/L dithiothreitol, pH 8.0) at a final volume of 100 μL were incubated at 25°C for 60 min. The reaction was stopped by the addition of 8 μL 0.25 mol/L EDTA and followed by the addition of 1 μL SYBR Green I (Beyotime Biotechnology, Shanghai, China). The fluorescence level was measured.
To evaluate the effects of temperature on the polymerase activity of POLD1, the DNA polymerase activity was measured at different temperatures ranging from 15°C to 45°C. To assess the thermostability of POLD1, the protein was incubated at 30°C, 35°C, 40°C, 45°C or 50°C for 20 min, 40 min, 60 min or 80 min. Then the residual activity of DNA polymerase was examined.
The effect of pH on DNA polymerase activity was evaluated at 25°C over a pH range of 5.0–10.0. Different buffers (40 mmol/L) were used: citric acid-Na2HPO4 (pH 5.0 to 6.0), phosphate buffer (pH 6.0 to 8.0) and boric acid-NaOH buffer (8.0 to 10.0). To evaluate the POLD1’s stability at different pH values, the protein was pre-incubated at a series of pH buffers (pH 5.0–10.0) for 20 min, 40 min, 60 min or 80 min. Subsequently the pH values of various reaction solutions were adjusted to pH 6.0 and the residual activity of POLD1 was examined.
The recombinant POLD1 was incubated with different chelating agent, reducing agent, protease inhibitor, detergents or metal ions at 25°C for 1 h. Subsequently the residual enzymatic activity was determined. The enzymatic activity without inhibitors or metal ions was defined as 100%. The protein inhibitors included ethylene diamine tetraacetic acid (EDTA), phenylmethanesulfonyl fluoride (PMSF), sodium dodecyl sulfate (SDS) or dithiothreitol (DTT) at a concentration of 1mmol/L and Triton-X 100 or Tween 20 at a concentration of 0.1% (w/v). To evaluate the influence of metal ions on the activity POLD1, the regular metal ions were chosen, such as Mn2+, Ca2+, Ba2+, Cu2+, K+ and Zn2+. The metal ions used were BaCl2·2H2O, CaCl2, CuCl·2H2O, KCL, ZnSO4·7H2O and MnCl2·4H2O at a final concentration of 10 mmol/L (Vasuvat et al., 2016; Vashishtha et al., 2016)
Kinetic characterizations of POLD1 were performed using different concentrations of dNTP (0.6 μmol/L, 0.8 μmol/L, 1.0 μmol/L, 1.2 μmol/L, 1.4 μmol/L, 1.6 μmol/L, 1.8 μmol/L and 2.0 μmol/L each) as the substrate. Kinetic parameters (Km and Vmax) were estimated by linear regression from Lineweaver- Burk plot.
All numerical data were analyzed by one-way ANOVA and expressed as mean ± standard deviation.
To characterize the DNA polymerase of deep-sea animals, POLD1 gene was cloned from vent shrimp R. exoculata. The results showed that a 2700-nt cDNA was obtained from R. exoculata (Fig. 1a). The cDNA contained an open reading frame (ORF), which encoded an 852-amino-acid protein (Fig. 1a) (GenBank accession no BankIt1979515 Seq1 KY412554). The POLD1 of R. exoculata contains a DNA polymerase type-B domain, three metal binding sites, and a zinc-finger domain (Fig. 1a). This 852-amino-acid protein shared high homology with the proteins in other animals. The phylogenetic analysis using neighbor-joining method showed that the R. exoculata POLD1 was close to that of Oratosquilla oratoria (Fig. 1b).

Recombinant shrimp POLD1 was expressed as a His tag fusion protein in bacteria E. coli and purified by Ni2+ affinity chromatography from soluble bacterial extracts. The results indicated that a band about 106 kDa corresponding to the POLD1-His tag fusion protein was observed in the induced bacteria containing the recombinant plasmid (Fig. 1c). No protein band was found at the same position in non-induced bacteria. SDS-PAGE showed that the recombinant POLD1 preparation contained a single 106 kDa polypeptide (Fig. 1c).
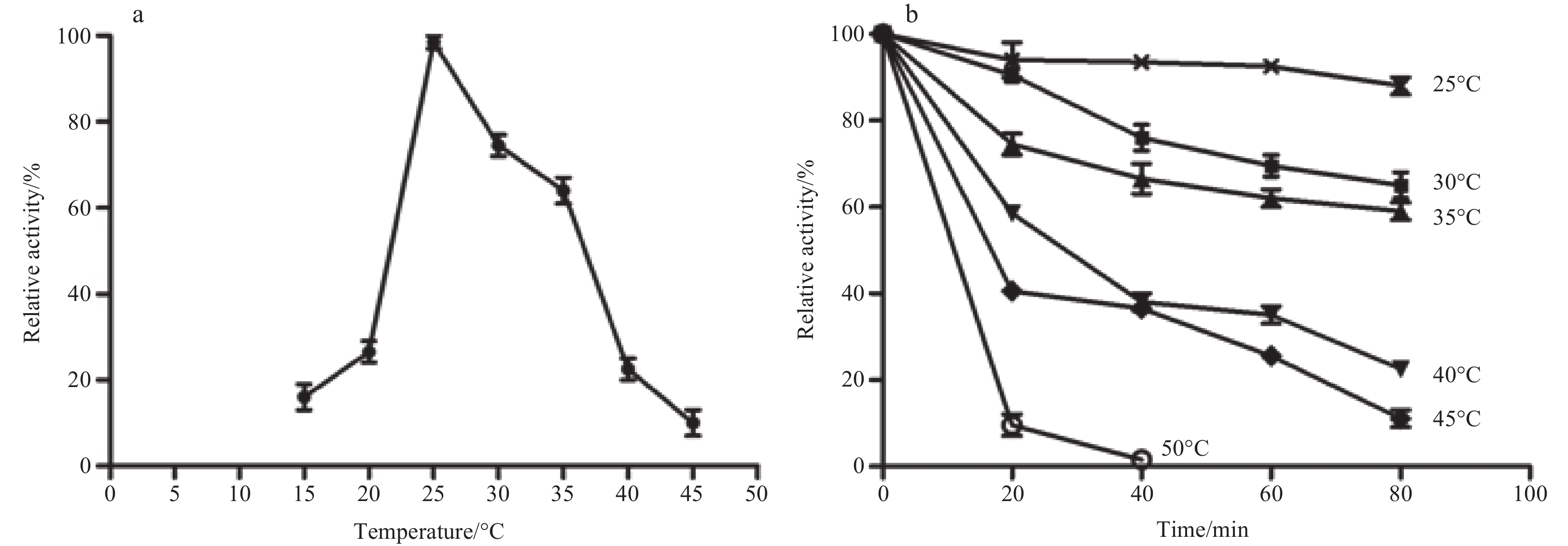
To evaluate the effects of temperature on the POLD1 polymerase activity of vent shrimp, the recombinant POLD1 polymerase activity was examined at different temperatures. The results indicated that POLD1 showed enzymatic activity over a range of 20°C to 40°C with an optimum temperature at 25°C corresponding to the R. exoculata’s living environment temperature (Fig. 2a). When the temperature was higher than 40°C, the polymerase activity was very low (Fig. 2a).

The thermostability assays revealed that the vent shrimp POLD1 was stable at 25°C for at least 80 min. When the temperature was increased to 30°C or 35°C, about 65% of the enzyme activity remained after 80 min. If the temperature rise to 45°C, the enzyme activity remained about 10% after 80 min. When the temperature was higher than 50°C, the polymerase activity decreased sharply (Fig. 2b).
To explore the optimum pH value of DNA polymerase activity, the enzymatic activity of vent shrimp POLD1 was monitored at different pH values. The results indicated that this enzyme was active over a wide range of 5.0 to 8.0, with an optimum at pH 6.0. The enzyme activity was only 39% and 26% of that of the optimum pH when the pH was 5 and 9 respectively, but remained 85% and 78% activity when the pH was 7 and 8, respectively (Fig. 3a). The recombinant polymerase showed a good tolerance when pH values maintain between 6 and 8. However, the polymerase activity decreased sharply at pH 5, 9, and 10 (Fig. 3b).

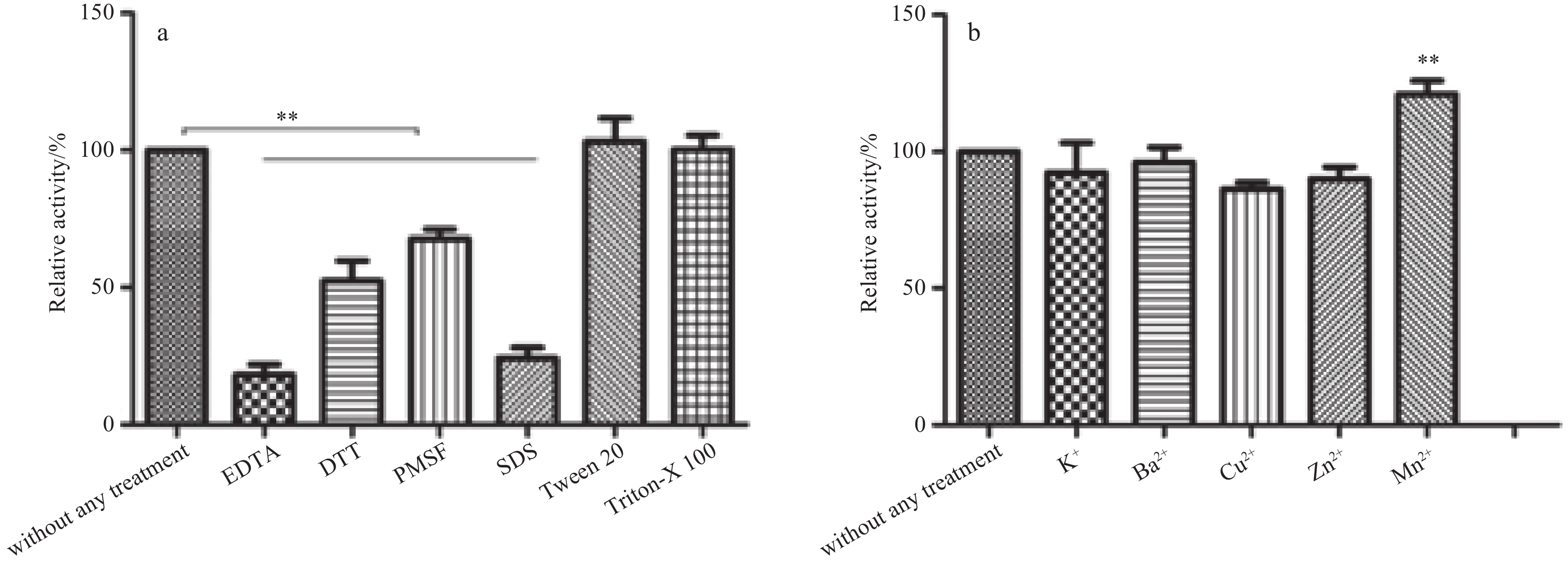
To evaluate the influence of inhibitors on the polymerase activity of POLD1, the recombinant POLD1 was incubated with different chelating agent, reducing agent, protease inhibitor and detergents, followed by the detection of residual activity of DNA polymerase. The results revealed that in the presence of EDTA and SDS, the enzyme activity was significantly decreased to 9.2% and 13.5%, respectively. In the present of protease inhibitor PMSF, the activity was 65% compare to that of the control. In the presence of detergents of Triton-X 100 and Tween 20, the polymerase activities of POLD1 were enhanced by 10.2% and 27.6%, respectively (Fig. 4a).

In order to assess the effects of metal ions on the polymerase activity, the recombinant enzyme was incubated with different metal ions and then the enzymatic activity was measured. The results indicated that the presence of K+, Ba2+, Cu2+ or Zn2+ has no obvious effect on the enzymatic activity, but the activity of the enzyme was significantly enhanced by Mn2+ (Fig. 4b).
To evaluate the affinity between enzyme and substrate, the kinetic parameters (Vmax and Km) of POLD1 were obtained from Lineweaver-Burke plots of specific activities at pH 6.5 and 25°C with different concentrations of dNTP. The results showed that Km value was 4.7 μmol/L and Vmax was 104.16 μmol/(L·min) (Fig. 5).

Replication of the leading and lagging strands of the eukaryotic nuclear genome involves multiple DNA polymerases. Among DNA polymerases, Polδ plays the most important role responsible for the synthesis of the lagging strand (Hübscher et al., 2002). To characterize the Polδ of deep-sea vent shrimp, the recombinant Polδ instead of the enzyme purified from vent shrimp tissues was employed in this study, because of the very limited number of vent shrimp from deep-sea hydrothermal vents.
In the present study, POLD1 of vent shrimp Rimicaris exoculata was successfully amplified and recombinantly expressed in E. coli. The results showed that the ORF of POLD1 of vent shrimp was 2700-nt. The deduced amino acid sequence (852 amino acids) shared high homology with the proteins in other animals. In eukaryotes, the POLD1 generally includes five domains, including two zinc-finger domains, the substrate binding domain, the center of the catalytic activity, the signal region of nucleus location, and the metal iron binding sites. Generally, the N-terminal of Polδ catalytic subunit contains no active domain (Baranovskiy et al., 2008; Biertümpfel et al., 2010). Within the DNA polymerase type-B delta subfamily catalytic domain, there is one metal-binding site, which is conserved in animals (Marchler-Bauer et al., 2011, 2015). In the present study, the recombinant POLD1 was shorter than the reported POLD1, due to the lack of N-terminal. The analysis indicated that POLD1 of vent shrimp contained three active domains, 3′-5′ exonuclease domain, DNA polymerase type-B delta subfamily catalytic domain and zinc-finger domain of DNA polymerase delta. In this context, the vent shrimp Polδ catalytic subunit could catalyze the synthesis of DNA in this study.
Divalent metal ions are essential components of DNA polymerases both for catalyzing nucleotide transfer reaction and base excision. All DNA polymerases require Mg2+ or Mn2+ for primer extension and for removal of incorrectly incorporated dNTPs (Vashishtha et al., 2016). Metal occupy sites A and B for DNA synthesis. A metal is in an octahedral complex composed of two highly conserved acidic residues, water molecules, and the 3-hydroxyl group of the primer. Its catalytic function is to reduce the hydroxyl pKa, making it a highly efficient nucleophilic cell, which can attack the phosphorus atoms of the incoming dNTPs. B metal helps to stabilize the transition state for nucleotide excision (Vashishtha et al., 2016). Substitution of Mg2+ with other divalent metal tend to reduce the fidelity of DNA replication (Vaisman et al., 2005). The results of this investigation showed that the chelate agent EDTA inhibited the DNA polymerase activity of POLD1, indicating that POLD1 requires divalent metal ions for primer extension. The fluids around DSHV where vent shrimp R. exoculata lives are enriched in metals, such as Na+, K+, Mg2+, Ca2+, Mn2+, Cu2+, Ba2+, Zn2+, and Co2+ (Hutnak et al., 2008). Metal Ba2+, Cu2+ or Zn2+ is not able to support the polymerase activity (Vashishtha et al., 2016). The results showed that the presence of K+, Ba2+, Cu2+ or Zn2+ has no obvious effect on the enzymatic activity, indicating that POLD1 of shrimp R. exoculata is stable under metal stress and its activity is not significantly affected by metal ions in the DSHV environment. Previous studies showed that Co2+ and Mn2+ could effectively replace Mg2+ but caused an increase in misincorporation (Seal et al., 1979; Hays and Berdis, 2002). The present results also showed the activity of the enzyme was significantly enhanced by Mn2+, indicating that Mn2+ may be a good substitute for Mg2+ in the terms of enzymatic activity. Due to limited opportunities and logistical difficulties in deep-sea sampling, the knowledge of metal bioaccumulation of deep-sea organisms and their relationships with environmental characteristics is quite scarce. Although there is no study on the concentration of metal ions in shrimp tissues from hydrothermal vents, studies from mussels show that despite quite different metals loads among the geochemical environment settings, Mn, Zn, and Cu concentrations varied over a smaller range across the sampling sites, implying biological regulation by deep-sea mussels for these elements (Zhou et al., 2020). In deep-sea hydrothermal vents, the vent shrimp R. exoculata are found to swarm around hydrothermal black smoker chimneys. This species maintains close proximity to the hydrothermal fluids, where steep thermal and chemical gradients are expected (Desbruyères et al., 2001). They inhabit the chimney wall on the gradient between hydrothermal fluid and cold oxygen environment seawater, and the temperature ranges from 3°C to 25°C (Le Bloa et al., 2017). Therefore, the species must evolve molecular mechanisms to cope with extreme temperature fluctuations. However, the understanding of the heat tolerance of this species and other deep-sea vent species is still limited due to the complexity of access to the vents and the difficulty of sampling and in vivo experiments on deep-sea animals under good physiological conditions. Deep-sea shrimp R. exoculata shows a higher thermal tolerance than the coastal shrimp Palaemonetes varians (Ravaux et al., 2019). This study showed that the polymerase activity of vent shrimp POLD1 was decreased when the temperature was higher than 30°C. The optimum temperature of vent shrimp POLD1 was 25°C, which was comparable to those of other marine shrimp DNA polymerases. The results also indicate that in a short period of time, such as 10 minutes, the effect of temperature on enzyme activity was not significant. This result has significant ecological implications, since this would mean that R. exoculata POLD1 is not affected by short temperature variations in its natural environment, which is an advantage in a thermal environment that is highly variable on short time scales. As reported, most types of DNA polymerases keep high activity under the condition of weak acid (Biertümpfel et al., 2010), which was consistent with the findings in this investigation.
The extreme conditions with sulfide, heavy metals, high pressure, and low dissolved oxygen in hydrothermal vents pose severe challenges to local inhabitants. Shrimps of the genus Rimicaris is one of the most studied hydrothermal crustaceans. However, the mechanism of their environmental adaptation and survival is unclear. A comparative transcriptome analysis of Rimicaris sp. was performed recently (Zhang et al., 2017). A large number of differentially expressed genes were found by comparing the transcriptome of the vent shrimp and the shrimp from the vent which were raised in the laboratory for 10 days. Among these differentially expressed genes, there are genes related to sulfide metabolism and anti-oxidative stress, which are likely involved in deep-sea survival. However, the adaptation strategies of deep-sea organisms to metals are not the same. A comparative transcriptome analysis of deep-sea mussels and their close relatives has shown that all mussels can adapt to different habitats, but different species may prefer to use different members of the same gene family (Zheng et al., 2017). In the present study, the activity of vent shrimp POLD1 was found stable under variousions and high concentrations, indicating that it was adapted to high salt concentration.
Considering the lack knowledge of the adaptation mechanism of deep-sea animals, the present results of vent shrimp POLD1 at least provide clues of potential adaptive molecular basis, and provide reference data for the future research of deep sea shrimp.
We are grateful to the 26th cruise of “China Ocean” for the shrimp R. exoculata used in this study.
| [1] |
Baranovskiy A G, Babayeva N D, Liston V G, et al. 2008. X-ray structure of the complex of regulatory subunits of human DNA polymerase delta. Cell Cycle, 7(19): 3026–3036. doi: 10.4161/cc.7.19.6720
|
| [2] |
Biertümpfel C, Zhao Ye, Kondo S, et al. 2010. Structure and mechanism of human DNA polymerase η. Nature, 465(7301): 1044–1048. doi: 10.1038/nature09196
|
| [3] |
Burgers P M J, Kunkel T A. 2017. Eukaryotic DNA replication fork. Annual Review of Biochemistry, 86: 417–438. doi: 10.1146/annurev-biochem-061516-044709
|
| [4] |
Desbruyères D, Biscoito M, Caprais J C, et al. 2001. Variations in deep-sea hydrothermal vent communities on the Mid-Atlantic Ridge near the Azores plateau. Deep-Sea Research Part I: Oceanographic Research Papers, 48(5): 1325–1346. doi: 10.1016/S0967-0637(00)00083-2
|
| [5] |
Fortune J M, Pavlov Y I, Welch C M, et al. 2005. Saccharomyces cerevisiae DNA polymerase δ: high fidelity for base substitutions but lower fidelity for single-and multi-base deletions. Journal of Biologial Chemistry, 280(33): 29980–29987. doi: 10.1074/jbc.M505236200
|
| [6] |
Haki G D, Rakshit S K. 2003. Developments in industrially important thermostable enzymes: a review. Bioresource Technology, 89(1): 17–34. doi: 10.1016/S0960-8524(03)00033-6
|
| [7] |
Hays H, Berdis A J. 2002. Manganese substantially alters the dynamics of translesion DNA synthesis. Biochemistry, 41(15): 4771–4778. doi: 10.1021/bi0120648
|
| [8] |
Hübscher U, Maga G, Spadari S. 2002. Eukaryotic DNA polymerases. Annual Review of Biochemistry, 71: 133–163. doi: 10.1146/annurev.biochem.71.090501.150041
|
| [9] |
Hutnak M, Fisher A T, Harris R, et al. 2008. Large heat and fluid fluxes driven through mid-plate outcrops on ocean crust. Nature Geoscience, 1: 611–614. doi: 10.1038/ngeo264
|
| [10] |
Jin Y H, Obert R, Burgers P M J, et al. 2001. The 3′→5′ exonuclease of DNA polymerase δ can substitute for the 5′ flap endonuclease Rad27/Fen1 in processing Okazaki fragments and preventing genome instability. Proceedings of the National Academy of Sciences of the United States of America, 98(9): 5122–5127. doi: 10.1073/pnas.091095198
|
| [11] |
Johnson R E, Prakash L, Prakash S. 2012. Pol31 and Pol32 subunits of yeast DNA polymerase δ are also essential subunits of DNA polymerase ζ. Proceedings of the National Academy of Sciences of the United States of America, 109(31): 12455–12460. doi: 10.1073/pnas.1206052109
|
| [12] |
Komaï T, Segonzac M. 2008. Taxonomic review of the hydrothermal vent shrimp genera Rimicaris Williams & Rona and Chorocaris Martin & Hessler (Crustacea: Decapoda: Caridea: Alvinocarididae). Journal of Shellfish Research, 27(1): 21–41. doi: 10.2983/0730-8000(2008)27[21:TROTHV]2.0.CO;2
|
| [13] |
Konn C, Charlou J L, Holm N G, et al. 2015. The production of methane, hydrogen, and organic compounds in ultramafic-hosted hydrothermal vents of the Mid-Atlantic Ridge. Astrobiology, 15(5): 381–399. doi: 10.1089/ast.2014.1198
|
| [14] |
Kumar S, Stecher G, Tamura K. 2016. MEGA7: Molecular evolutionary genetics analysis version 7. 0 for bigger datasets. Molecular Biology and Evolution, 33(7): 1870–1874. doi: 10.1093/molbev/msw054
|
| [15] |
Kunkel T A, Burgers P M J. 2017. Arranging eukaryotic nuclear DNA polymerases for replication: Specific interactions with accessory proteins arrange Pols α, δ, and ϵ in the replisome for leading-strand and lagging-strand DNA replication. Bioessays, 39(8): 1700070. doi: 10.1002/bies.201700070
|
| [16] |
Le Bloa S, Durand L, Cueff-Gauchard V, et al. 2017. Highlighting of quorum sensing lux genes and their expression in the hydrothermal vent shrimp Rimicaris exoculata ectosymbiontic community. Possible use as biogeographic markers. PLoS ONE, 12(3): e0174338
|
| [17] |
Maga G, Villani G, Tillement V, et al. 2001. Okazaki fragment processing: modulation of the strand displacement activity of DNA polymerase δ by the concerted action of replication protein A, proliferating cell nuclear antigen, and flap endonuclease-1. Proceedings of the National Academy of Sciences of the United States of America, 98(25): 14298–14303. doi: 10.1073/pnas.251193198
|
| [18] |
Marchler-Bauer A, Derbyshire M K, Gonzales N R, et al. 2015. CDD: NCBI’s conserved domain database. Nucleic Acids Research, 43: D222–D226. doi: 10.1093/nar/gku1221
|
| [19] |
Marchler-Bauer A, Lu Shennan, Anderson J B, et al. 2011. CDD: a conserved domain database for the functional annotation of proteins. Nucleic Acids Research, 39: D225–D229. doi: 10.1093/nar/gkq1189
|
| [20] |
Prindle M J, Loeb L A. 2012. DNA polymerase delta in DNA replication and genome maintenance. Environmental and Molecular Mutagenesis, 53(9): 666–682. doi: 10.1002/em.21745
|
| [21] |
Ravaux J, Cottin D, Chertemps T, et al. 2009. Hydrothermal vent shrimps display low expression of the heat-inducible hsp70 gene in nature. Marine Ecology Progress Series, 396: 153–156. doi: 10.3354/meps08293
|
| [22] |
Ravaux J, Léger N, Hamel G, et al. 2019. Assessing a species thermal tolerance through a multiparameter approach: the case study of the deep-sea hydrothermal vent shrimp Rimicaris exoculata. Cell Stress and Chaperones, 24: 647–659. doi: 10.1007/s12192-019-01003-0
|
| [23] |
Rayner E, van Gool I C, Palles C, et al. 2016. A panoply of errors: polymerase proofreading domain mutations in cancer. Nature Review Cancer, 16: 71–81. doi: 10.1038/nrc.2015.12
|
| [24] |
Seal G, Shearman C W, Loeb L A. 1979. On the fidelity of DNA replication: studies with human placenta DNA polymerases. Journal of Biological Chemistry, 254(12): 5229–5237. doi: 10.1016/S0021-9258(18)50583-4
|
| [25] |
Swan M K, Johnson R E, Prakash L, et al. 2009. Structural basis of high-fidelity DNA synthesis by yeast DNA polymerase δ. Nature Structural & Moecularl Biology, 16: 979–986
|
| [26] |
Tveit H, Kristensen T. 2001. Fluorescence-based DNA polymerase assay. Analytical Biochemistry, 289(1): 96–98. doi: 10.1006/abio.2000.4903
|
| [27] |
Vaisman A, Ling Hong, Woodgate R, et al. 2005. Fidelity of Dpo4: effect of metal ions, nucleotide selection and pyrophosphorolysis. EMBO Journal, 24: 2957–2967. doi: 10.1038/sj.emboj.7600786
|
| [28] |
Vashishtha A K, Wang Jimin, Konigsberg W H. 2016. Different divalent cations alter the kinetics and fidelity of DNA polymerases. Journal of Biological Chemistry, 291(40): 20869–20875. doi: 10.1074/jbc.R116.742494
|
| [29] |
Vasuvat J, Montree A, Moonsom S, et al. 2016. Biochemical and functional characterization of Plasmodium falciparum DNA polymerase δ. Malaria Journal, 15: 116. doi: 10.1186/s12936-016-1166-0
|
| [30] |
Weedon M N, Ellard S, Prindle M J, et al. 2013. An in-frame deletion at the polymerase active site of POLD1 causes a multisystem disorder with lipodystrophy. Nature Genetics, 45: 947–950. doi: 10.1038/ng.2670
|
| [31] |
Williams A B, Rona P A. 1986. Two new caridean shrimps (bresiliidae) from a hydrothermal field on the Mid-Atlantic Ridge. Journal of Crustacean Biology, 6(3): 446–462. doi: 10.1163/193724086X00299
|
| [32] |
Zhang Jian, Sun Qinglei, Luan Zhendong, et al. 2017. Comparative transcriptome analysis of Rimicaris sp. reveals novel molecular features associated with survival in deep-sea hydrothermal vent. Scientific Reports, 7: 2000
|
| [33] |
Zheng Ping, Wang Minxiao, Li Chaolun, et al. 2017. Insights into deep-sea adaptations and host-symbiont interactions: a comparative transcriptome study on Bathymodiolus mussels and their coastal relatives. Molecular Ecology, 26(19): 5133–5148. doi: 10.1111/mec.14160
|
| [34] |
Zhou Li, Cao Lei, Wang Xiaocheng, et al. 2020. Metal adaptation strategies of deep-sea Bathymodiolus mussels from a cold seep and three hydrothermal vents in the West Pacific. Science of the Total Environment, 707: 136046. doi: 10.1016/j.scitotenv.2019.136046
|


Figures(6)

Supported by:
Beijing Renhe Information Technology Co. Ltd

Wenlin Wu, Hongyun Li, Tiantian Ma, Xiaobo Zhang. Characterization of DNA polymerase δ from deep-sea hydrothermal vent shrimp Rimicaris exoculata[J]. Acta Oceanologica Sinica, 2021, 40(8): 168-175. doi: 10.1007/s13131-021-1823-1

 DownLoad:
DownLoad:

 DownLoad:
DownLoad: