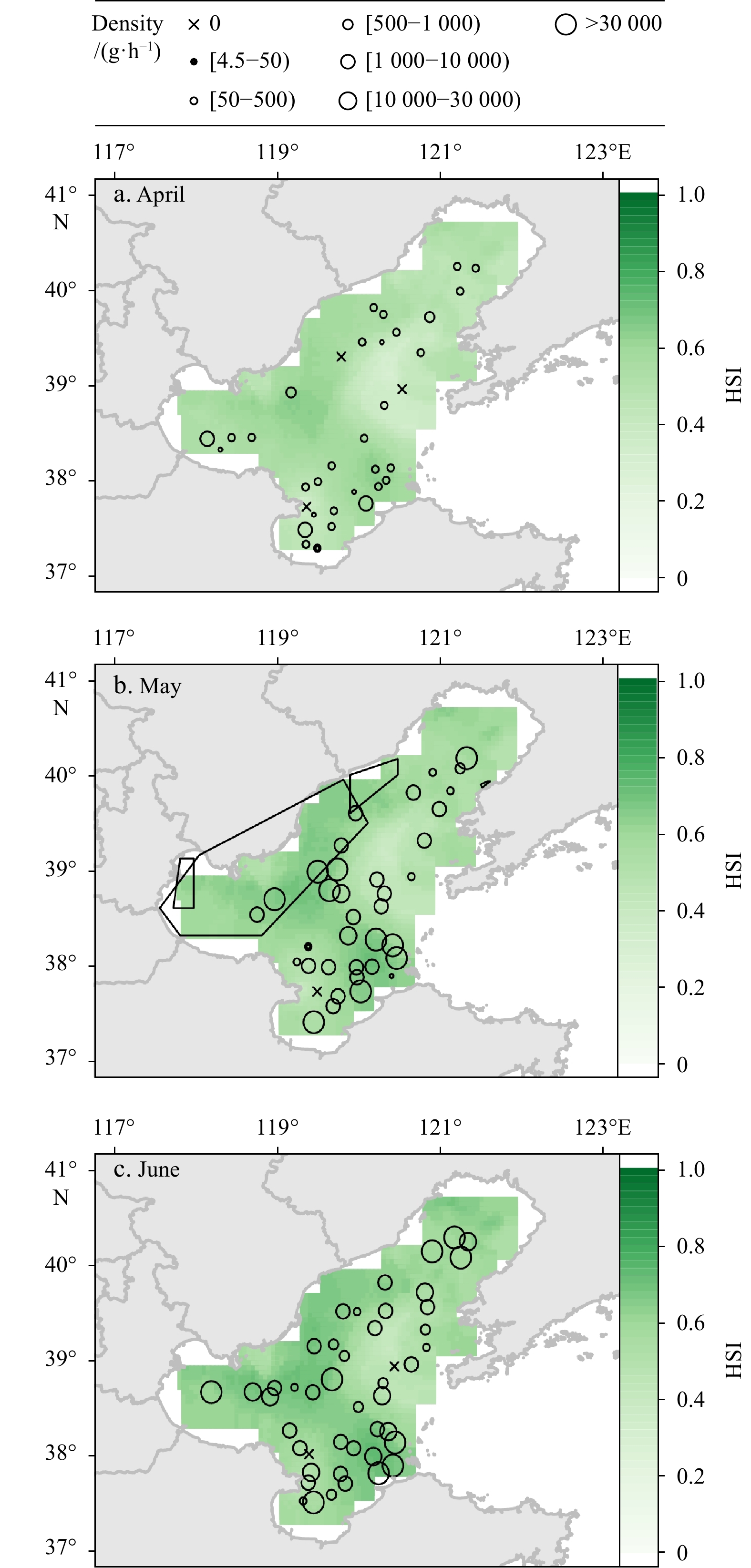
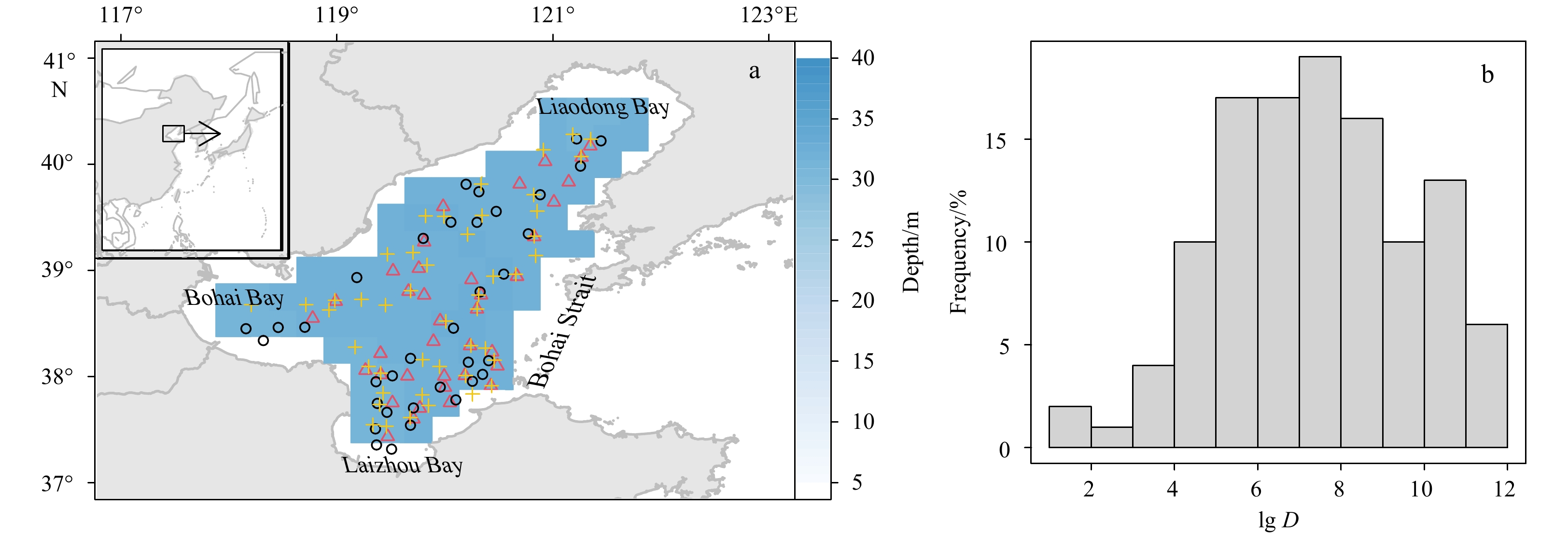
| Citation: | Lisha Guan, Xianshi Jin, Tao Yang, Xiujuan Shan. Ensemble habitat suitability modeling of stomatopods with Oratosquilla oratoria as an example[J]. Acta Oceanologica Sinica, 2023, 42(4): 93-102. doi: 10.1007/s13131-022-2051-z

|
|
Abo-Hashesh A T, Madkour F F, Sallam W S, et al. 2020. Molecular identification and phylogenetic relationship of Erugosquilla massavensis (Kossmann, 1880) from the Mediterranean Sea, Egypt. Egyptian Journal of Aquatic Biology and Fisheries, 24(6): 533–545. doi: 10.21608/ejabf.2020.117576
|
|
Antony P J, Dhanya S, Lyla P S, et al. 2010. Ecological role of stomatopods (mantis shrimps) and potential impacts of trawling in a marine ecosystem of the southeast coast of India. Ecological Modelling, 221(21): 2604–2614. doi: 10.1016/j.ecolmodel.2010.07.017
|
|
Cerasoli F, Besnard A, Marchand M A, et al. 2021. Determinants of habitat suitability models transferability across geographically disjunct populations: Insights from Vipera ursinii ursinii. Ecology and Evolution, 11(9): 3991–4011. doi: 10.1002/ece3.7294
|
|
Chen Tung-Yun, Hwang Gwo-Wen, Mayfield A B, et al. 2017. The relationship between intertidal soil composition and fiddler crab burrow depth. Ecological Engineering, 100: 256–260. doi: 10.1016/j.ecoleng.2016.12.011
|
|
Chen Tung-Yun, Hwang Gwo-Wen, Mayfield A B, et al. 2019. The development of habitat suitability models for fiddler crabs residing in subtropical tidal flats. Ocean & Coastal Management, 182: 104931
|
|
Covich A P, Palmer M A, Crowl T A. 1999. The role of benthic invertebrate species in freshwater ecosystems: Zoobenthic species influence energy flows and nutrient cycling. Bioscience, 49(2): 119–127. doi: 10.2307/1313537
|
|
DeVries M S, Stock B C, Christy J H, et al. 2016. Specialized morphology corresponds to a generalist diet: linking form and function in smashing mantis shrimp crustaceans. Oecologia, 182(2): 429–442. doi: 10.1007/s00442-016-3667-5
|
|
Ding Qi, Shan Xiujuan, Jin Xianshi, et al. 2020. Research on utilization conflicts of fishery resources and catch allocation methods in the Bohai Sea, China. Fisheries Research, 225: 105477. doi: 10.1016/j.fishres.2019.105477
|
|
Franklin J. 2009. Mapping Species Distributions: Spatial Inference and Prediction. Cambridge: Cambridge University Press, 181–187
|
|
Georgian S E, Anderson O F, Rowden A A. 2019. Ensemble habitat suitability modeling of vulnerable marine ecosystem indicator taxa to inform deep-sea fisheries management in the South Pacific Ocean. Fisheries Research, 211: 256–274. doi: 10.1016/j.fishres.2018.11.020
|
|
Gianguzza P, Insacco G, Zava B, et al. 2019. Much can change in a year: The massawan mantis shrimp, Erugosquilla massavensis (Kossmann, 1880) in sicily, Italy. BioInvasions Records, 8(1): 108–112. doi: 10.3391/bir.2019.8.1.11
|
|
Gollini I, Lu Binbin, Charlton M, et al. 2015. GWmodel: An R package for exploring spatial heterogeneity using geographically weighted models. Journal of Statistical Software, 63(17): 1–50
|
|
Heikkinen R K, Marmion M, Luoto M. 2012. Does the interpolation accuracy of species distribution models come at the expense of transferability?. Ecography, 35(3): 276–288
|
|
Kim S, Kim H, Bae H, et al. 2017. Growth and reproduction of the Japanese mantis shrimp, Oratosquilla oratoria (De Haan 1844) in the coastal area of Tongyeong, Korea. Ocean Science Journal, 52(2): 257–265. doi: 10.1007/s12601-017-0027-2
|
|
Kodama K, Shimizu T, Yamakawa T, et al. 2006. Changes in reproductive patterns in relation to decline in stock abundance of the Japanese mantis shrimp Oratosquilla oratoria in Tokyo Bay. Fisheries Science, 72(3): 568–577. doi: 10.1111/j.1444-2906.2006.01185.x
|
|
Krumbein W C. 1934. Size Frequency distributions of sediments. Journal of Sedimentary Research, 4(2): 65–77
|
|
Laverock B, Gilbert J A, Tait K, et al. 2011. Bioturbation: Impact on the marine nitrogen cycle. Biochemical Society Transactions, 39(1): 315–320. doi: 10.1042/BST0390315
|
|
Li Bai, Cao Jie, Guan Lisha, et al. 2018. Estimating spatial non-stationary environmental effects on the distribution of species: a case study from American lobster in the Gulf of Maine. ICES Journal of Marine Science, 75(4): 1473–1482. doi: 10.1093/icesjms/fsy024
|
|
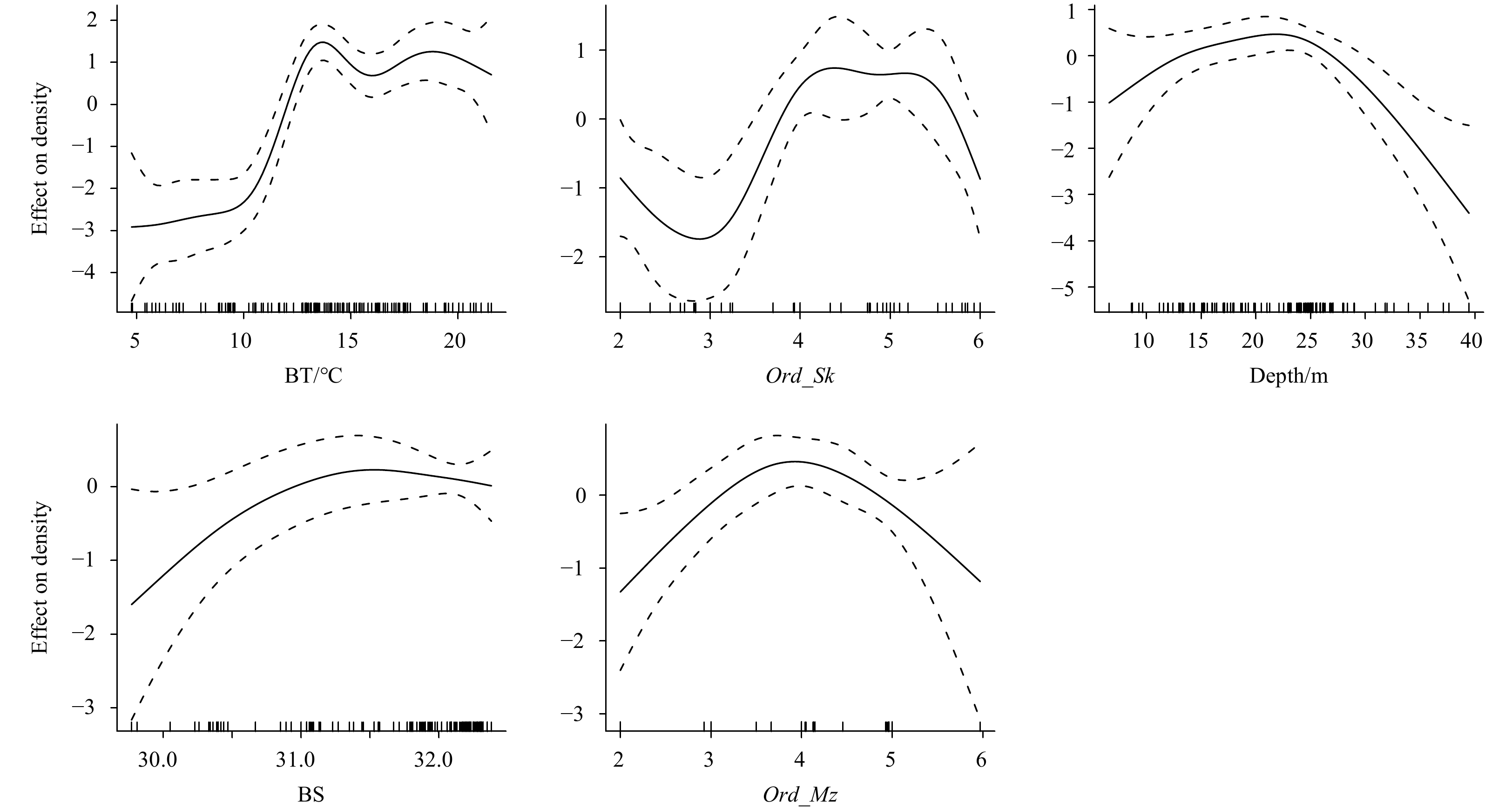
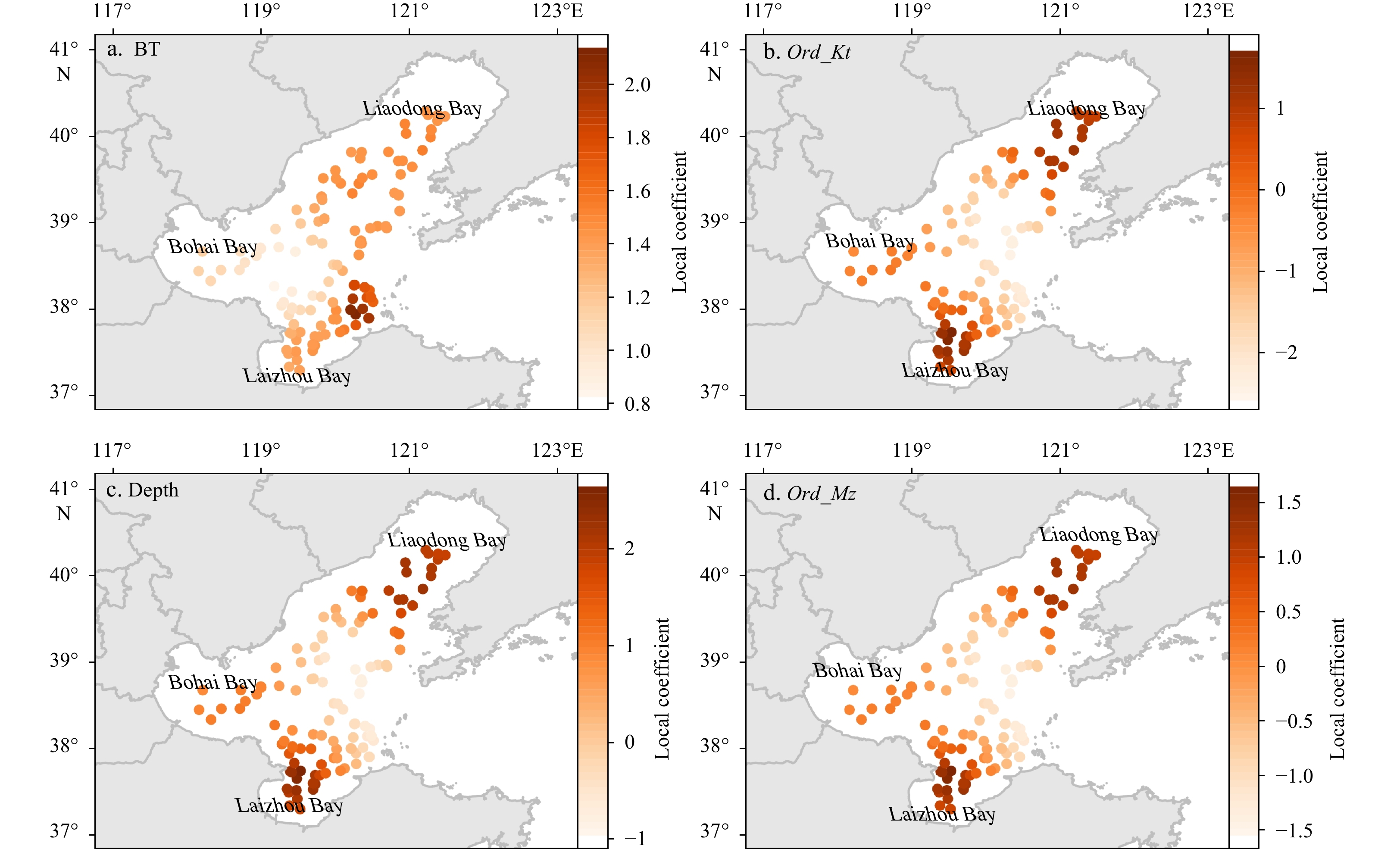
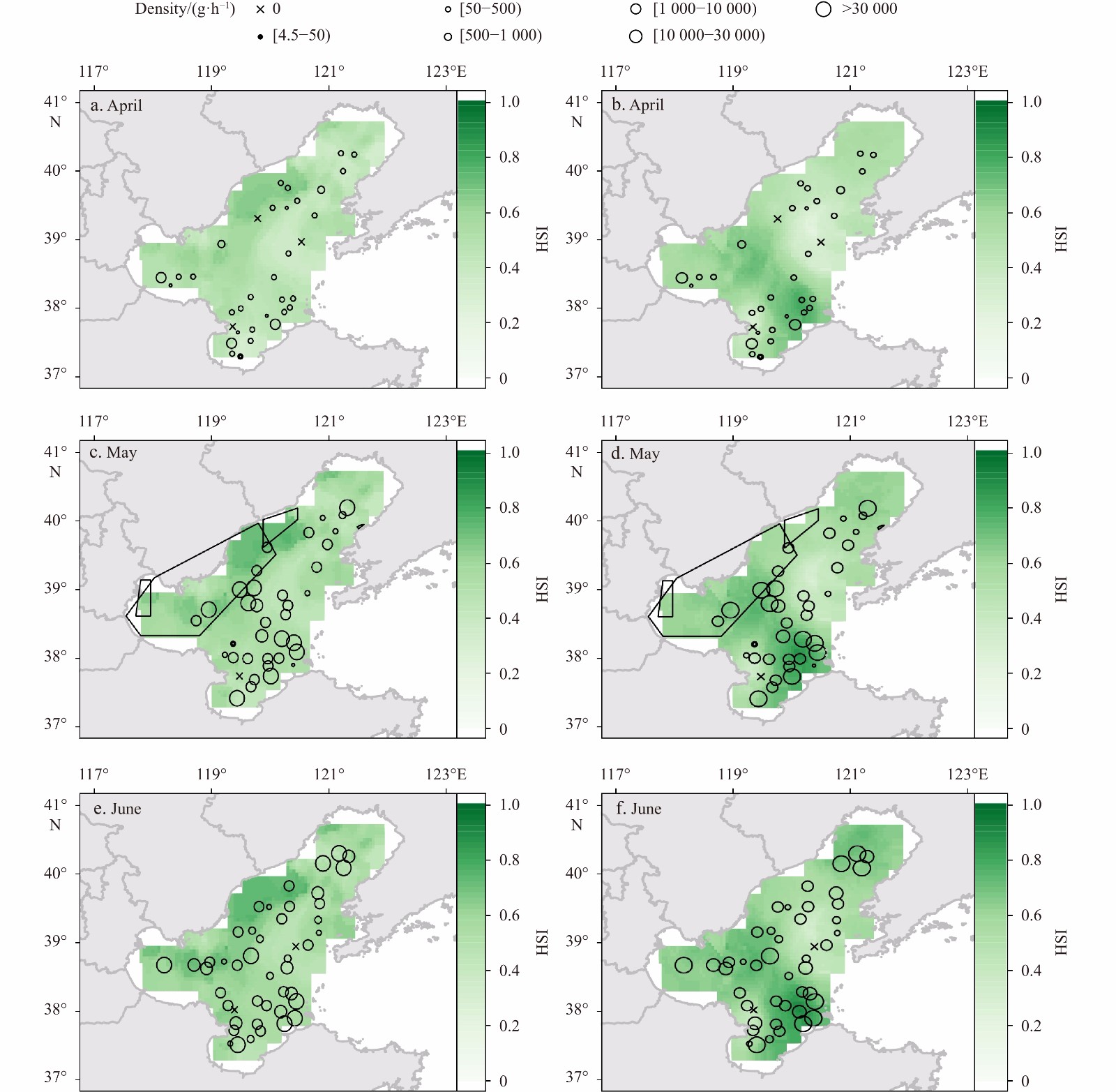
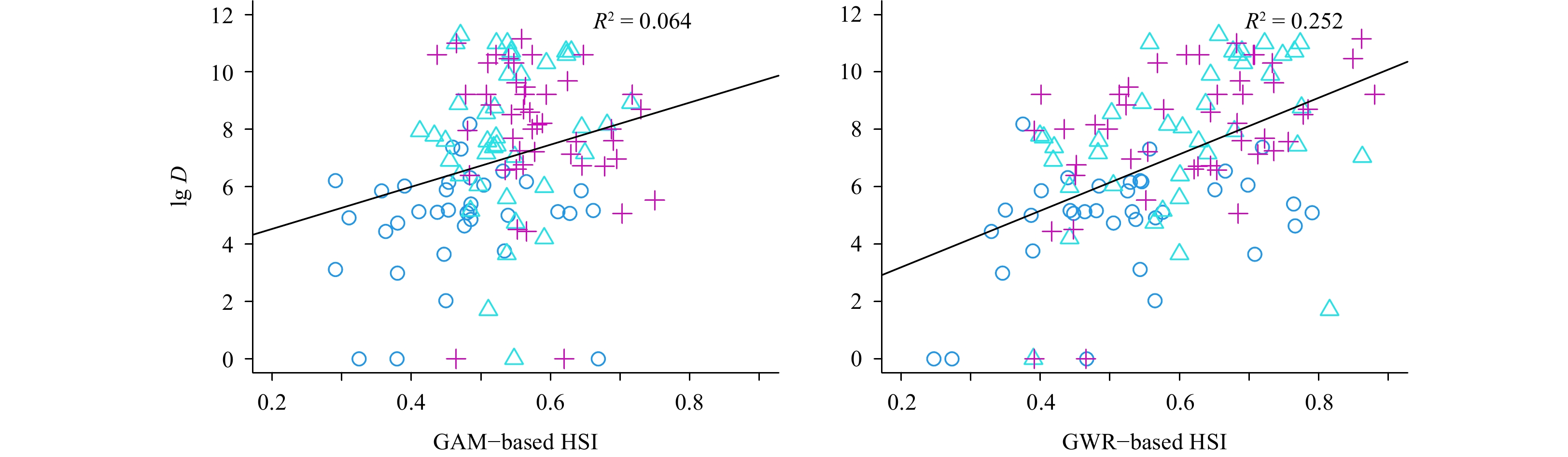
Li Mingkun, Zhang Chongliang, Xu Binduo, et al. 2020. A comparison of GAM and GWR in modelling spatial distribution of Japanese mantis shrimp (Oratosquilla oratoria) in coastal waters. Estuarine, Coastal and Shelf Science, 244: 106928
|
|
Liu Haiying, Wang Gui’e, Wang Xiuli. 2009. Genetic diversity analysis of mantis shrimp Oratosquilla oratoria from Dalian coast. Journal of Dalian Fisheries University (in Chinese), 24(4): 350–353
|
|
Lou Fangrui, Gao Tianxiang, Han Zhiqiang. 2019. Effect of salinity fluctuation on the transcriptome of the Japanese mantis shrimp Oratosquilla oratoria. International Journal of Biological Macromolecules, 140: 1202–1213. doi: 10.1016/j.ijbiomac.2019.08.223
|
|
Lu Binbin, Charlton M, Harris P, et al. 2014. Geographically weighted regression with a non-Euclidean distance metric: A case study using hedonic house price data. International Journal of Geographical Information Science, 28(4): 660–681. doi: 10.1080/13658816.2013.865739
|
|
Lui K K Y, Leung P T Y, Ng W C, et al. 2010. Genetic variation of Oratosquilla oratoria (Crustacea: Stomatopoda) across Hong Kong waters elucidated by mitochondrial DNA control region sequences. Journal of the Marine Biological Association of the United Kingdom, 90(3): 623–631. doi: 10.1017/S0025315409990841
|
|
Luo Chongxin, Lin Lei, Shi Jie, et al. 2021. Seasonal variations in the water residence time in the Bohai Sea using 3D hydrodynamic model study and the adjoint method. Ocean Dynamics, 71(2): 157–173. doi: 10.1007/s10236-020-01438-5
|
|
Manning R B. 1971. Keys to the species of Oratosquilla (Crustacea, Stomatopoda), with descriptions of two new species. Smithsonian Contributions to Zoology, 1: 1–16
|
|
Maynou F, Abelló P, Sartor P. 2004. A review of the fisheries biology of the mantis shrimp, Squilla mantis (L., 1758) (Stomatopoda, Squillidae) in the Mediterranean. Crustaceana, 77(9): 1081–1099. doi: 10.1163/1568540042900295
|
|
Nakajima M, Kodama K, Horiguchi T, et al. 2010. Impacts of shifts in spawning seasonality and size at maturation on the population growth of mantis shrimp in Tokyo Bay. Marine Ecology Progress Series, 418: 179–188. doi: 10.3354/meps08824
|
|
Ng Yingpei. 2013. The Ecology of Stomatopods in Matang waters with emphasis on Miyakea nepa and Oratosquillina perpensa [dissertation]. Kuala Lumpur: University of Malaya
|
|
Qiao Huijie, Feng Xiao, Escobar L E, et al. 2019. An evaluation of transferability of ecological niche models. Ecography, 42(3): 521–534. doi: 10.1111/ecog.03986
|
|
Ragheb E, Akel E S H K, Rizkalla S I. 2019. Analyses of the non-target catch from the Egyptian Mediterranean trawlers, off Port Said. The Egyptian Journal of Aquatic Research, 45(3): 239–246. doi: 10.1016/j.ejar.2019.07.003
|
|
Reaka M L. 1980. Geographic range, life history patterns, and body size in a guild of coral-dwelling mantis shrimps. Evolution, 34(5): 1019–1030
|
|
Reaka M L, Rodgers P J, Kudla A U. 2008. Patterns of biodiversity and endemism on Indo-West Pacific coral reefs. Proceedings of the National Academy of Sciences of the United States of America, 105(S1): 11474–11481
|
|
Robert K, Jones D O B, Roberts J M, et al. 2016. Improving predictive mapping of deep-water habitats: Considering multiple model outputs and ensemble techniques. Deep-Sea Research Part I: Oceanographic Research Papers, 113: 80–89. doi: 10.1016/j.dsr.2016.04.008
|
|
Tanaka K, Chen Y. 2015. Spatiotemporal variability of suitable habitat for american lobster (Homarus Americanus) in Long Island sound. Journal of Shellfish Research, 34(2): 531–543. doi: 10.2983/035.034.0238
|
|
Tao L S R, Lui K K Y, Lau E T C, et al. 2018. Trawl ban in a heavily exploited marine environment: Responses in population dynamics of four stomatopod species. Scientific Reports, 8(1): 17876. doi: 10.1038/s41598-018-35804-7
|
|
Wang Lei, Qiu Shengrao, Liu Shude, et al. 2020. Morphological difference analysis on three different stocks of Oratosquilla oratoria in the Bohai Sea and the Yellow Sea. Marine Fisheries (in Chinese), 42(6): 672–686
|
|
Wilson J. 2017. Learning, adaptation, and the complexity of human and natural interactions in the ocean. Ecology and Society, 22(2): 43. doi: 10.5751/ES-09356-220243
|
|
Wu Qiang, Guan Lisha, Shan Xiujuan, et al. 2019. Decadal variations in the community status of economically important invertebrates in the Bohai Sea. Acta Oceanologica Sinica, 38(10): 60–66. doi: 10.1007/s13131-019-1488-1
|
|
Xue Ying, Guan Lisha, Tanaka K, et al. 2017. Evaluating effects of rescaling and weighting data on habitat suitability modeling. Fisheries Research, 188: 84–94. doi: 10.1016/j.fishres.2016.12.001
|
|
Yang Jiming. 2001. A study on food and trophic levels of Bohai Sea Invertebrates. Modern Fisheries Information (in Chinese), 16(9): 8–16
|
|
Yang Mei, Li Xinzheng. 2018. Population genetic structure of the mantis shrimp Oratosquilla oratoria (Crustacea: Squillidae) in the Yellow Sea and East China Sea. Journal of Oceanology and Limnology, 36(3): 905–912. doi: 10.1007/s00343-018-7030-z
|
|
Yuan Ping, Wang Houjie, Wu Xiao, et al. 2020. Grain-size distribution of surface sediments in the Bohai Sea and the northern Yellow Sea: sediment supply and hydrodynamics. Journal of Ocean University of China, 19(3): 589–600. doi: 10.1007/s11802-020-4221-y
|
|
Zhang Yang, Han Zhiqiang, Gao Tianxiang, et al. 2018. Genetic structure analysis of mantis shrimp Oratosquilla oratoria based on mitochondrial DNA control region sequence. Genes and Genomics, 40(9): 1001–1009. doi: 10.1007/s13258-018-0707-z
|
|
Zhang Yunlei, Yu Huaming, Yu Haiqing, et al. 2020. Optimization of environmental variables in habitat suitability modeling for mantis shrimp Oratosquilla oratoria in the Haizhou Bay and adjacent waters. Acta Oceanologica Sinica, 39(6): 36–47. doi: 10.1007/s13131-020-1546-8
|
|
Zhao Wenju, Cao Taohong, Dou Pinxin, et al. 2019. Effect of various concentrations of superabsorbent polymers on soil particle-size distribution and evaporation with sand mulching. Scientific Reports, 9(1): 3511. doi: 10.1038/s41598-019-39412-x
|
| 1. | Anglu Shen, Shufei Gao, Christopher M. Heggerud, et al. Fluctuation of growth and photosynthetic characteristics in Prorocentrum shikokuense under phosphorus limitation: Evidence from field and laboratory. Ecological Modelling, 2023, 479: 110310. doi:10.1016/j.ecolmodel.2023.110310 | |
| 2. | Tiantian Chen, Yun Liu, Sha Xu, et al. Variation of Amoebophrya community during bloom of Prorocentrum donghaiense Lu in coastal waters of the East China Sea. Estuarine, Coastal and Shelf Science, 2020, 243: 106887. doi:10.1016/j.ecss.2020.106887 | |
| 3. | Yong Zhang, Jin-Zhu Su, Yu-Ping Su, et al. Spatial Distribution of Phytoplankton Community Composition and Their Correlations with Environmental Drivers in Taiwan Strait of Southeast China. Diversity, 2020, 12(11): 433. doi:10.3390/d12110433 | |
| 4. | Yaqun Zhang, Xin Lin, Tangcheng Li, et al. High throughput sequencing of 18S rRNA and its gene to characterize a Prorocentrum shikokuense (Dinophyceae) bloom. Harmful Algae, 2020, 94: 101809. doi:10.1016/j.hal.2020.101809 | |
| 5. | Hao Zhang, Jiuling Liu, Yanbin He, et al. Quantitative proteomics reveals the key molecular events occurring at different cell cycle phases of the in situ blooming dinoflagellate cells. Science of The Total Environment, 2019, 676: 62. doi:10.1016/j.scitotenv.2019.04.216 | |
| 6. | Lixia Shang, Zhangxi Hu, Yingzhong Tang. Identification of dissolved and particulate carbonyl compounds produced by marine harmful algal bloom species. Journal of Oceanology and Limnology, 2019, 37(5): 1566. doi:10.1007/s00343-019-8199-5 | |
| 7. | Yaqun Zhang, Xin Lin, Xinguo Shi, et al. Metatranscriptomic Signatures Associated With Phytoplankton Regime Shift From Diatom Dominance to a Dinoflagellate Bloom. Frontiers in Microbiology, 2019, 10 doi:10.3389/fmicb.2019.00590 |
Figures(7) / Tables(2)

Supported by:
Beijing Renhe Information Technology Co. Ltd

SUN Ke, QIU Zhongfeng, HE Yijun, FAN Wei, WEI Zexun. Vertical development of a Prorocentrum donghaiense bloom in the coastal waters of the East China Sea:coupled biophysical numerical modeling[J]. Acta Oceanologica Sinica, 2017, 36(6): 23-33. doi: 10.1007/s13131-016-0965-z





 DownLoad:
DownLoad: