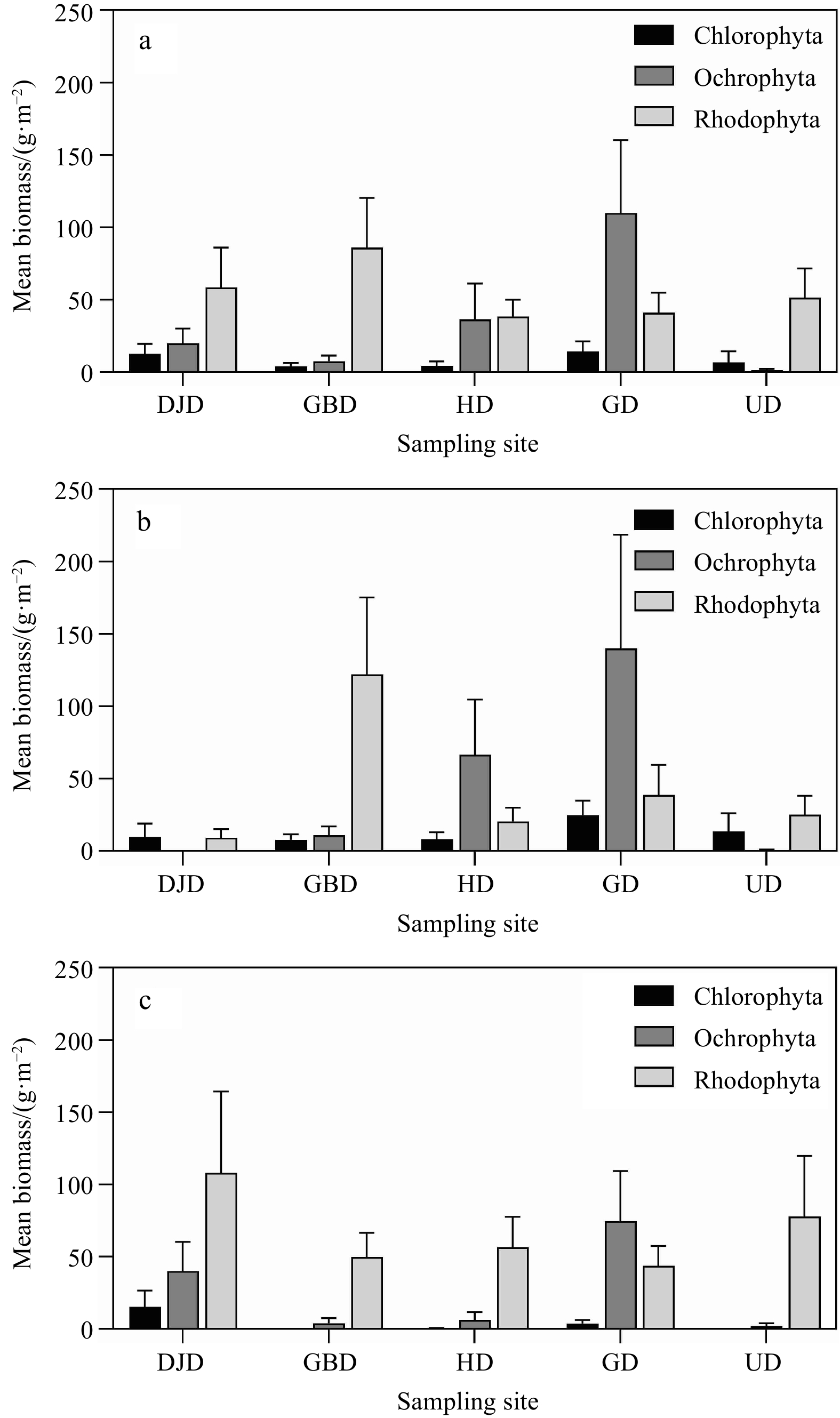
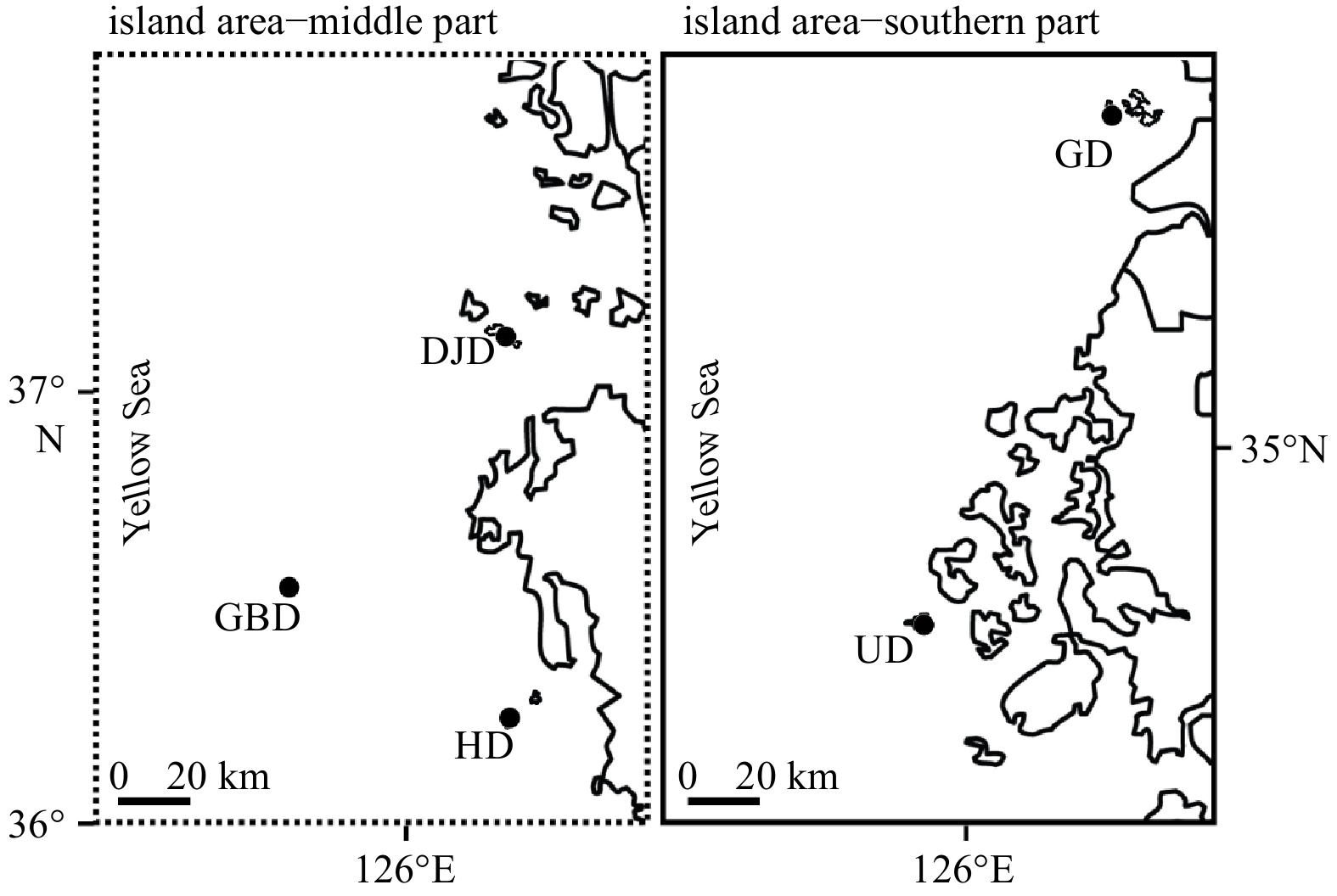
| Citation: | Su Jin Han, Jae-Gil Jang, Hyun-Jung Kim, Tae-Ho Seo, Joo Myun Park. Ecological evaluation of marine macroalgal communities on five islands of Korea in the Yellow Sea[J]. Acta Oceanologica Sinica, 2023, 42(6): 49-56. doi: 10.1007/s13131-022-2089-y

|
|
Ahn J K, Yoo K D, Oh J C, et al. 2017. Species composition and vertical distribution of marine algal communities at the Taean Peninsula of the west coast of Korea. Korean Journal of Fisheries and Aquatic Sciences, 50(1): 55–64. doi: 10.5657/KFAS.2017.0055
|
|
Akrong M O, Anning A K, Addico G N D, et al. 2021. Spatio-temporal variations in seaweed diversity and abundance of selected coastal areas in Ghana. Regional Studies in Marine Science, 44: 101719. doi: 10.1016/j.rsma.2021.101719
|
|
Baek J M, Hwang M S, Lee J W, et al. 2007. The macroalgal community of Bagryoungdo Island in Korea. Algae (in Korean), 22(2): 117–123. doi: 10.4490/ALGAE.2007.22.2.117
|
|
Best E P H, Buzzelli C P, Bartell S M, et al. 2001. Modeling submersed macrophyte growth in relation to underwater light climate: modeling approaches and application potential. Hydrobiologia, 444(1): 43–70. doi: 10.1023/A:1017564632427
|
|
Caldeira A Q, De Paula J C, Reis R P, et al. 2017. Structural and functional losses in macroalgal assemblages in a southeastern Brazilian bay over more than a decade. Ecological Indicators, 75: 242–248. doi: 10.1016/j.ecolind.2016.12.029
|
|
Chabane K, Bahbah L, Seridi H. 2018. Ecological quality status of the Algiers coastal waters by using macroalgae assemblages as bioindicators (Algeria, Mediterranean Sea). Mediterranean Marine Science, 19(2): 305–315. doi: 10.12681/mms.15951
|
|
Cheney D P. 1977. A new and improved ratio for comparing seaweed floras. Journal of Phycology, 13(S1): 12
|
|
Cho T O, Boo S M. 1996. Seasonal changes of marine plants in Oeyondo Island on the Yellow Sea. Algae (in Korean), 11(3): 285–293
|
|
Choi D S, Kim K Y, Lee W J, et al. 1994. Marine algal flora and community structure of Uido Island, west-southern coast of Korea. Korean Journal of Environmental Biology (in Korean), 12: 65–75
|
|
Choi H G, Lee K H, Wan X Q, et al. 2008. Temporal variations in seaweed biomass in Korean coasts: Woejodo and Jusamdo, Jeonbuk. Algae (in Korean), 23(4): 335–342. doi: 10.4490/ALGAE.2008.23.4.335
|
|
Choi C G, Rho H S. 2010. Marine algal community of Ulsan, on the eastern coast of Korea. Korean Journal of Fisheries and Aquatic Science, 43(3): 246–253. doi: 10.5657/kfas.2010.43.3.246
|
|
Clarke K R, Gorley R N. 2006. PRIMER V6: User Manual/Tutorial. Plymouth, UK: PRIMER-E Ltd
|
|
Cohen C, Fowler J. 1990. Practical Statistics for Field Biology. Maidenhead: Open University Press
|
|
Edwards M S, Connell S D. 2012. Competition, a major factor structuring seaweed communities. In: Wiencke C, Bischof K, eds. Seaweed Biology. Berlin: Springer, 135–156
|
|
Guiry M D, Guiry G M. 2021. AlgaeBase. http://www.Algaebase.org[2021-10-08]
|
|
Heo J S, Park S K, Yoo H I, et al. 2011. Macroalgal community structure on the rocky shores of Ongdo, Jusamdo and Woejodo islands of the Yellow Sea. Fisheries and Aquatic Science, 14(4): 389–397. doi: 10.5657/FAS.2011.0389
|
|
Hwang E K, Park C S, Sohn C H, et al. 1996. Analysis of functional form groups in macroalgal community of Yonggwang vicinity, western coast of Korea. Korean Journal of Fisheries and Aquatic Sciences (in Korean), 29(1): 97–106
|
|
Juanes J A, Guinda X, Puente A, et al. 2008. Macroalgae, a suitable indicator of the ecological status of coastal rocky communities in the NE Atlantic. Ecological Indicator, 8(4): 351–359. doi: 10.1016/j.ecolind.2007.04.005
|
|
Jung R H, Hwang D W, Kim Y G, et al. 2010. Temporal variations in the sedimentation rate and benthic environment of intertidal surface sediments around Byeonsan Peninsula, Korea. Korean Journal of Fisheries and Aquatic Science (in Korean), 43(6): 723–734. doi: 10.5657/kfas.2010.43.6.723
|
|
Jung S W, Rho H S, Choi C G. 2020. Characteristics and ecosystem changes of marine algal communities in Wangdol-cho on the east coast of Korea. Ocean Science Journal, 55(4): 549–562. doi: 10.1007/s12601-020-0034-6
|
|
Kang J W. 1966. On the geographical distribution of marine algae in Korea. Bulletin of Pusan Fisheries College, 7: 1–125
|
|
Kim H S, Boo S M, Lee I K, et al. 2013a. National List of Species of Korea: Marine Algae. Incheon: National Institute of Biological Resources
|
|
Kim B Y, Kim W S, Choi H G. 2010. Seasonal variability of seaweed biomass along the vertical shore gradients of Nachido and Odo islands, the Yellow Sea, Korea. Fisheries and Aquatic Sciences, 13(4): 324–331. doi: 10.5657/fas.2010.13.4.324
|
|
Kim J H, Ko Y D, Kim Y S, et al. 2011. Marine algal flora and community structure of Gogunsan islands outside the Saemangeum Dike. Korean Journal of Environment and Ecology (in Korean), 25(2): 156–165
|
|
Kim Y S, Yang E A, Nam K W. 2013b. Benthic marine algal flora and community structure of Eocheongdo in western coast of Korea. Korean Journal of Environment and Ecology (in Korean), 27(6): 655–665. doi: 10.13047/KJEE.2013.27.6.655
|
|
Koh C H, Oh S H. 1992. Distribution pattern of macroalgae in the eastern Yellow Sea, Korea. The Korean Journal of Phycology, 7: 139–146
|
|
Kong D, Kim A R. 2015. Analysis on the relationship between biological indices and survey area of benthic macroinvertebrates using mathematical model. Journal of Korean Society on Water Environment (in Korean), 31(6): 610–618. doi: 10.15681/KSWE.2015.31.6.610
|
|
Lee W J, Hwang M S, Baek J M, et al. 2007. Primary survey on algal community of Gyounggi Bay for restoration. Algae (in Korean), 22(3): 201–207. doi: 10.4490/ALGAE.2007.22.3.201
|
|
Liang Zhourui, Wang Feijiu, Sun Xiutao, et al. 2014. Reproductive biology of Sargassum thunbergii (Fucales, Phaeophyceae). American Journal of Plant Sciences, 5(17): 2574–2581. doi: 10.4236/ajps.2014.517271
|
|
Littler M M, Littler D S. 1984. Relationships between macroalgal functional form groups and substrata stability in a subtropical rocky-intertidal system. Journal of Experimental Marine Biology and Ecology, 74(1): 13–34. doi: 10.1016/0022-0981(84)90035-2
|
|
Liu Jinlin, Tong Yichao, Xia Jing, et al. 2022. Ulva macroalgae within local aquaculture ponds along the estuary of Dagu River, Jiaozhou Bay, Qingdao. Marine Pollution Bulletin, 174: 113243. doi: 10.1016/j.marpolbul.2021.113243
|
|
Liu Jinlin, Xia Jing, Zhuang Minmin, et al. 2021a. Golden seaweed tides accumulated in Pyropia aquaculture areas are becoming a normal phenomenon in the Yellow Sea of China. Science of the Total Environment, 774: 145726. doi: 10.1016/j.scitotenv.2021.145726
|
|
Liu Jinlin, Xia Jing, Zhuang Minmin, et al. 2021b. Controlling the source of green tides in the Yellow Sea: NaClO treatment of Ulva attached on Pyropia aquaculture rafts. Aquaculture, 535: 736378. doi: 10.1016/j.aquaculture.2021.736378
|
|
Margalef R. 1958. Information theory in ecology. General Systems, 3: 36–71
|
|
McNaughton S J. 1967. Relationships among functional properties of Californian Grassland. Nature, 216(5111): 168–169. doi: 10.1038/216168b0
|
|
Oh J C, Choi H G, Kim C D, et al. 2016. Ecological evaluation of marine national parks based on seaweed community index. Korean Journal of Fisheries and Aquatic Sciences (in Korean), 49(3): 385–392. doi: 10.5657/KFAS.2016.0385
|
|
Oh B G, Lee J W, Lee H B. 2005. Summer marine algal vegetation of uninhabited islands in Sinangun, southwestern coast. Algae (in Korean), 20(1): 53–59. doi: 10.4490/ALGAE.2005.20.1.053
|
|
Orfanidis S, Panayotidis P, Stamatis N. 2001. Ecological evaluation of transitional and coastal waters: A marine benthic macrophytes-based model. Mediterranean Marine Science, 2(2): 45–66. doi: 10.12681/mms.266
|
|
Orfanidis S, Panayotidis P, Stamatis N. 2003. An insight to the ecological evaluation index (EEI). Ecological Indicators, 3(1): 27–33. doi: 10.1016/S1470-160X(03)00008-6
|
|
Orfanidis S, Panayotidis P, Ugland K. 2011. Ecological evaluation index continuous formula (EEI-c) application: a step forward for functional groups, the formula and reference condition values. Mediterranean Marine Science, 12(1): 199–232. doi: 10.12681/mms.60
|
|
Padilla D K, Allen B J. 2000. Paradigm lost: reconsidering functional form and group hypotheses in marine ecology. Journal of Experimental Marine Biology and Ecology, 250(1–2): 207–221,
|
|
Park C S, Hwang E K. 2011. An investigation of the relationship between sediment particles size and the development of green algal mats (Ulva prolifera) on the intertidal flats of Muan, Korea. Journal of Applied Phycology, 23(3): 515–522. doi: 10.1007/s10811-010-9620-9
|
|
Park C S, Park K Y, Hwang E K. 2013. Benthic algal flora in a man-made artificial beach in the Hwawon Resort Complex, southwestern coast of Korea. Korean Journal of Environmental Biology (in Korean), 31(2): 78–86. doi: 10.11626/KJEB.2013.31.2.078
|
|
Park Y K, Seo K S, Choi C K. 2006. Environmental Biology (in Korean). Seoul: Daehakseolim
|
|
Park C S, Wee M Y, Hwang E K. 2007. Summer algal flora of uninhabited islands in Dochodo, southwestern coast of Korea. Algae (in Korean), 22(4): 305–311. doi: 10.4490/ALGAE.2007.22.4.305
|
|
Pinedo S, García M, Satta M P, et al. 2007. Rocky-shore communities as indicators of water quality: a case study in the northwestern Mediterranean. Marine Pollution Bulletin, 55(1–6): 126–135,
|
|
Roleda M Y, Morris J N, Mcgraw C M, et al. 2012. Ocean acidification and seaweed reproduction: Increased CO2 ameliorates the negative effect of lowered pH on meiospore germination in the giant kelp Macrocystis pyrifera (Laminariales, Phaeophyceae). Global Change Biology, 18(3): 854–864. doi: 10.1111/j.1365-2486.2011.02594.x
|
|
Romdoni T A, Ristiani A, Meinita M D N, et al. 2018. Seaweed species composition, abundance and diversity in Drini and Kondang Merak Beach, Java. E3S Web of Conferences, 47: 03006. doi: 10.1051/e3sconf/20184703006
|
|
Russell B D, Thompson J A I, Falkenberg L J, et al. 2009. Synergistic effects of climate change and local stressors: CO2 and nutrient-driven change in subtidal rocky habitats. Global Change Biology, 15(9): 2153–2162. doi: 10.1111/j.1365-2486.2009.01886.x
|
|
Saito Y, Atobe S. 1970. Phytosociological study of intertidal marine algae: I. Usujiri Benten-Jima, Hokkaido. Bulletin of the Faculty of Fisheries Hokkaido University, 21(2): 37–69
|
|
Scherner F, Horta P A, De Oliveira E C, et al. 2013. Coastal urbanization leads to remarkable seaweed species loss and community shifts along the SW Atlantic. Marine Pollution Bulletin, 76(1–2): 106–115,
|
|
Song W O, Jin J Y, Chae J W, et al. 2003. A review of measures against environmental impact of suspended sediments generated by coastal development works. Ocean and Polar Research (in Korean), 25(spc3): 409–416. doi: 10.4217/OPR.2003.25.spc3.409
|
|
Sun Yuqing, Yao Lulu, Liu Jinlin, et al. 2022. Prevention strategies for green tides at source in the southern Yellow Sea. Marine Pollution Bulletin, 178: 113646. doi: 10.1016/j.marpolbul.2022.113646
|
|
Vale C G, Arenas F, Barreiro R, et al. 2021. Understanding the local drivers of beta-diversity patterns under climate change: The case of seaweed communities in Galicia, North West of the Iberian Peninsula. Diversity and Distributions, 27(9): 1696–1705. doi: 10.1111/ddi.13361
|
|
Wells E, Wilkinson M, Wood P, et al. 2007. The use of macroalgal species richness and composition on intertidal rocky seashores in the assessment of ecological quality under the European water framework directive. Marine Pollution Bulletin, 55(1–6): 151–161,
|
|
Whitaker S G, Smith J R, Murray S N. 2010. Reestablishment of the southern California rocky intertidal brown alga, Silvetia compressa: An experimental investigation of techniques and abiotic and biotic factors that affect restoration success. Restoration Ecology, 18: 18–26. doi: 10.1111/j.1526-100X.2010.00717.x
|
|
Yang E A, Kim Y S. 2009. Summer marine algal flora of Sipidongpado, located in the middle western coast of Korea. Korean Journal of Nature Conservation (in Korean), 3(1): 1–5
|
|
Yoo J S, Kim Y H. 2003. Community dynamics of the benthic marine algae in Hakampo, the western coast of Korea. Korean Journal of Environmental Biology, 21(4): 428–438
|
|
Yoo H I, Lee J H, Lee K H, et al. 2007. Summer marine algal floras and community structures in Taean Peninsula, Korea. Korean Journal of Fisheries and Aquatic Sciences (in Korean), 40(4): 210–219. doi: 10.5657/kfas.2007.40.4.210
|
|
Yoon M Y, Boo S M. 1991. Flora and zonation of marine plants at the littoral area of Sapsido Island on the Yellow Sea of Korea. Korean Journal of Phycology (in Korean), 6: 145–156
|
Figures(2) / Tables(6)

Supported by:
Beijing Renhe Information Technology Co. Ltd

BI Haibo, HUANG Haijun, SU Qiao, YAN Liwen, LIU Yanxia, XU Xiuli. An Arctic sea ice thickness variability revealed from satellite altimetric measurements[J]. Acta Oceanologica Sinica, 2014, 33(11): 134-140. doi: 10.1007/s13131-014-0562-y





 DownLoad:
DownLoad: