Yueming Wu, Yongjun Wei, Juan Severino Pino Ibánhez, Danqing Sun, Kaiqi Lu, Qianqian Wang, Jian Li, Shan Jiang. Microbiota of a seepage face at the mouth of a subterranean estuary: diversity, distribution and substrate dependence[J]. Acta Oceanologica Sinica, 2023, 42(8): 147-157. doi: 10.1007/s13131-023-2257-8
Citation:
Yueming Wu, Yongjun Wei, Juan Severino Pino Ibánhez, Danqing Sun, Kaiqi Lu, Qianqian Wang, Jian Li, Shan Jiang. Microbiota of a seepage face at the mouth of a subterranean estuary: diversity, distribution and substrate dependence[J]. Acta Oceanologica Sinica, 2023, 42(8): 147-157. doi: 10.1007/s13131-023-2257-8
Yueming Wu, Yongjun Wei, Juan Severino Pino Ibánhez, Danqing Sun, Kaiqi Lu, Qianqian Wang, Jian Li, Shan Jiang. Microbiota of a seepage face at the mouth of a subterranean estuary: diversity, distribution and substrate dependence[J]. Acta Oceanologica Sinica, 2023, 42(8): 147-157. doi: 10.1007/s13131-023-2257-8
Citation:
Yueming Wu, Yongjun Wei, Juan Severino Pino Ibánhez, Danqing Sun, Kaiqi Lu, Qianqian Wang, Jian Li, Shan Jiang. Microbiota of a seepage face at the mouth of a subterranean estuary: diversity, distribution and substrate dependence[J]. Acta Oceanologica Sinica, 2023, 42(8): 147-157. doi: 10.1007/s13131-023-2257-8
Subterranean estuaries, i.e., the mixing zone between terrestrial groundwater and recirculated seawater, host a wide range of microbiota. Here, field campaigns were conducted at the mouth of the subterranean estuary at the Sanggou Bay (Shandong Province, China) over four consecutive seasons at a seepage face (0−20 cm depth). The diversity of benthic microbiome was characterized via 16S rRNA gene sequencing and metagenomics, combined with physic-chemical parameters, e.g., organic carbon, total nitrogen and sulfate contents in sediments. During spring, the dominant species were assigned to the phylum Proteobacteria. Important opportunistic species was assigned to Acidobacteria, Actinobacteria and Bacteroidetes. The key components were identified to be species of the genera Pseudoalteromonas, Colwellia and Sphingobium, indicating the involvement of sediment microbiota in the degradation of sedimentary organic carbon, particularly that of pelagic origin, e.g., phytoplankton detritus and bivalve pseudo-feces. During spring, the microbial community was statistically similar along the depth profiles and among the three sampled stations. Similar spatial distributions were obtained in the remaining seasons. By contrast, the dominant species assemblages varied significantly among seasons, with key genera being Thioprofundum and Nitrosopumilus during summer and autumn and Thioprofundum and Ilumatobacter during winter. Network analysis revealed a seasonal shift in benthic nitrogen and sulfur metabolism associated with these variations in microbial community composition. Overall, our findings suggested that macro elements derived from pelagic inputs, particularly detrital phytoplankton, shaped the microbial community compositions at the seepage face, resulting in significant seasonal variations, while the influence of terrestrial materials transported by groundwater on the sediment microbiota at the seepage face found to be minor.
Submarine groundwater discharge (SGD), i.e., the outflow from permeable coastal sediments or karst conducts into the receiving oceans, is a commonly overlooked pathway for transporting terrestrial nutrients into coastal seawater (Moore, 2010; Santos et al., 2021). Recent estimates based on radioactive tracers suggested that this transport pathway represented the largest source of terrestrial solutes to the global ocean (Cho et al., 2018). However, accurate estimating of solute fluxes through this pathway is challenging owing to the inherent uncertainties associated with our limited understanding of the environmental capacity of subterranean estuaries (STEs) in modulating SGD-driven nutrient fluxes. As the mixing zone between terrestrial groundwater and seawater, STEs are recognized as important conductors that host a wide range of biogeochemical reactions, significantly altering the porewater macro element (C, N, S and P) concentrations and chemical species prior to discharge (Jiang et al., 2021b; Rocha et al., 2021). However, the reaction capability of STEs on solutes is highly variable, primarily due to the co-occurrence of different biogeochemical reactions with conflicting effects on target solutes, such as nitrification and denitrification/biological assimilation (Rocha et al., 2022; Jiang et al., 2023). To achieve a precise quantification of chemical element loading into the coast from SGD, several research projects funded by governments have focused on investigating the spatial and temporal variations of solute concentrations in STEs and aiming to explore the dynamic linkage between environmental variables, such as temperature, salinity, organic carbon supply, and solute concentration changes (Reckhardt et al., 2015; Linkhorst et al., 2017; Calvo-Martin et al., 2021; Jiang et al., 2023).
Benthic reactions in modulating solute transport through SGD to the coast have been associated with highly diverse microbiota (Jiang et al., 2020; Ruiz-González et al., 2022). Nevertheless, compared to research on solute distribution and reactivity within STEs, our knowledge on the responsible microbiota is significantly limited (Archana et al., 2021). Till 2021, only 12 research publications describing distribution and diversity of microorganisms were identified, covering sporadic sites in China, US and Europe (Archana et al., 2021). More importantly, the spatial distribution and seasonal variation of STE microbiota, along with their elemental drivers, and benthic microbial function, remain largely underexplored. The lack of comprehensive understanding of the tight linkage between biogeochemical reactions and benthic microbiota in STEs hampers the exploration of the solute modulation capability in STEs, especially in different seasons, and therefore precise estimation of nutrient fluxes associated with SGD.
In the present study, field surveys were conducted during four consecutive seasons in the Sanggou Bay, Shandong Province, China to unveil the compositions, spatial and temporal distributions, as well as environmental drivers of the microbial community of this STE. The specific objectives of this study were: (1) to characterize spatial and temporal variations of the benthic microbiota at the SGD seepage face; (2) to evaluate the links between benthic microbial communities and metabolism; (3) to identify the pelagic and terrestrial drivers, particularly element supply, of the distribution of benthic microbial diversity. Based on these investigations, we aim to provide insights into the microbial dynamics within the STE and its interactions with the surrounding environment.
2.
Materials and methods
2.1
Study sites and sample collection
The Sanggou Bay (location outlined in Fig. S1; temperate monsoon climate) is as a significant aquaculture hub for the cultivation of kelp (Laminaria laminaria), Pacific oyster (Crassostrea gigas), and scallops (Chlamys farreri). The bay water frequently exhibits high phytoplankton biomass, with diatoms being the dominant species (Yuan et al., 2014). The Guhe River presents the highest discharge rate of all, ranging from 1.7×108 m3/a to 2.3×108 m3/a (Wang et al., 2014). Apart from the Guhe River, SGD, fed from the adjacent coastal aquifers, is also important for the transport of terrestrial solutes (Jiang et al., 2023), particularly from the highly permeable sediments of the inner boundary of the bay (Jiang et al., 2020). During ebbing tides, a significant amount of low-salinity groundwater seeps from the beach into the bay water (Jiang et al., 2020). The annual precipitation is around 820 mm.
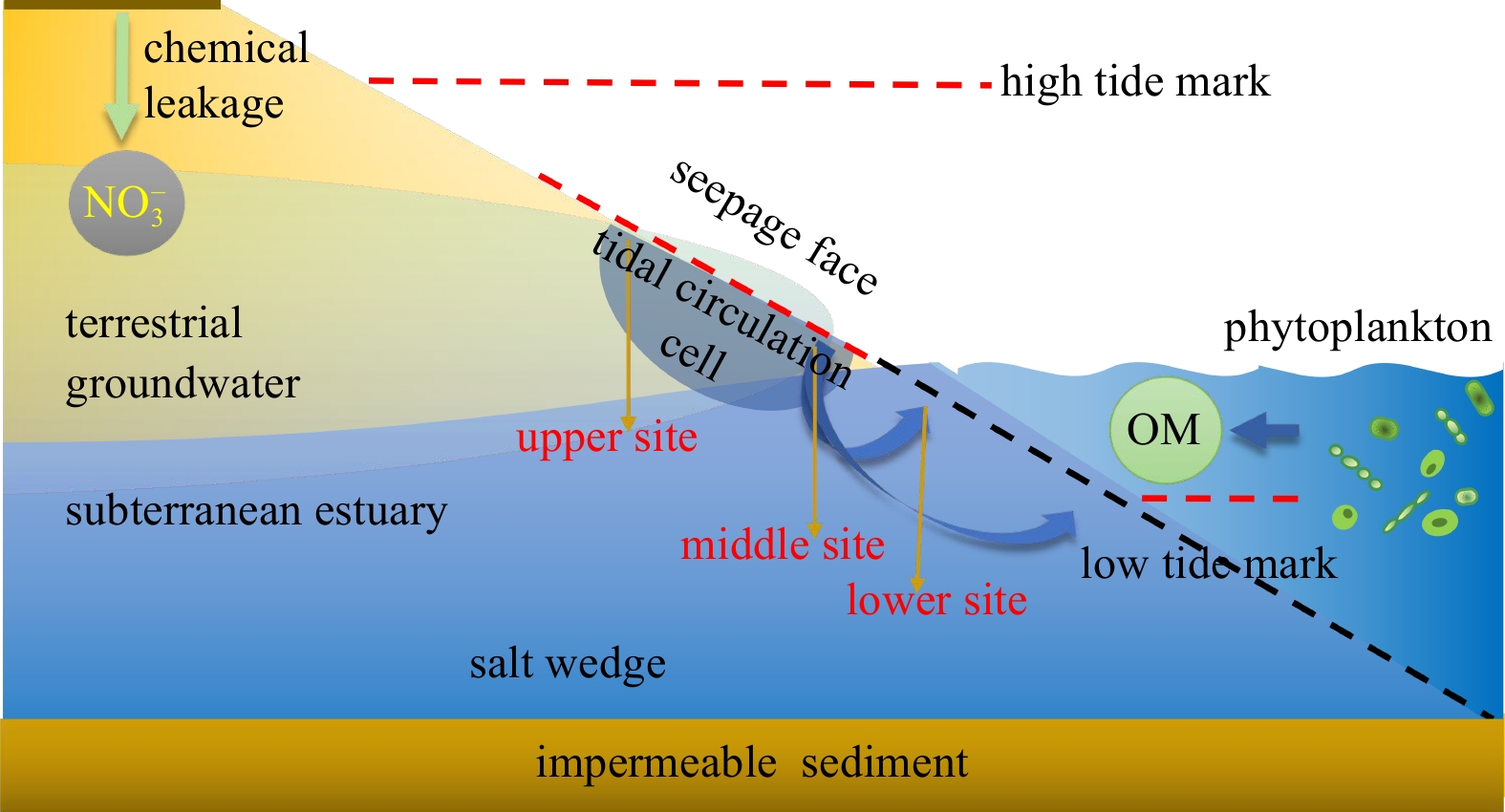
Four seasonal field surveys (spring, April 2018; summer, July 2018; autumn, October 2018; winter, February 2019) were conducted at the seepage face of the inner boundary of the Sanggou Bay (37°08'25"N, 122°29'07"E). Seepage face is the outer regime of STE, receiving great quantity of pelagic compounds compared with terrestrial materials (Jiang et al., 2023). During each sampling campaign, three sampling stations were chosen at the upper, middle, and lower intertidal flat in the seepage face, as depicted in Fig. 1. Sediment samples were collected from each station of the seepage face using polymethyl methacrylate sediment corers, from depths of 0−20 cm. The sediment column was rapidly sliced into five sections (1−4 cm, 5−8 cm, 9−12 cm, 13−16 cm, and 17−20 cm depth) and stored in sterilized bags at −20℃ prior to laboratory analyses. The first centimeter was discarded to remove the accumulation of shell debris and plastic pieces.
Figure
1.
Sketch of subterranean estuary (STE) and seepage face in coastal zones. This figure highlights the three sampling sites (upper, middle and lower tidal stations) and major organic matter (OM) sources for STE. This figure was modified from Jiang et al. (2021a).
After collection, sediment subsamples were dried at 50℃ in an oven until constant weight was achieved. Subsequently, algae and bivalve debris were removed from the sediment subsamples. Sediment grain size was determined using a Coulter LS 100Q instrument (Coulter Company, USA), and particles larger than 63 μm were identified as the sandy fraction (Wu et al., 2019). Total organic carbon (TOC) was quantified using a CHNOS Elemental Analyzer (Vario EL III, Germany), after exposure of the sample to 2 mol/L hydrochloric acid flumes overnight to remove inorganic carbon. Total nitrogen (TN) and total sulfur (TS) in the sediment were quantified using a CHNS Cube (Wei et al., 2022). Reactive phosphorus (RP) content in the sediment was extracted using a 0.5 mol/L potassium sulfate solution and quantified in a flow injection system (SAN plus, SKALAR Analytical B.V., The Netherlands). Additionally, the colored dissolved organic matter (CDOM) content in the extracted solution was analyzed using a spectrophotometer (TU-1901, PERSEE®, China). CDOM contents in the tested water samples were estimated based on the absorption coefficient at 355 nm wavelength (a355) (Zhang et al., 2021).
The microbial DNA was extracted from the fresh wet sediments using DNeasy PowerSoil kit (Qiagen, Germany) for microbial community analysis. Except samples from spring (only top and bottom layers in each boundary), all sediment samples from the remaining seasons were included in the microbial analysis. The 16S rRNA amplicon was applied to reveal microbial structure in the sediment according to previous study (Liang et al., 2019), and the raw data was deposited in the Sequence Read Archive (SRA) database. In order to give insight into the microbial metabolism of the nutrients, the metagenomic sequencing strategy following Liang et al. (2021) was applied for both summer and autumn sediment samples. In particular, microbial DNA extracted from these STE sediments were subject to high-throughput sequencing, and the obtained reads were assembled and analyzed (Wei et al., 2020). The genes from the assembled contigs were annotated with NCBI-NR, KEGG, and STRING-COG databases. The KEGG and COG functional categories were calculated based on the quantity of EGTs assigned to the specific KO or COG. The metagenomic sequences were submitted to the SRA database.
2.3
Mathematic analyses for microbial information
The obtained microbial data (fastq files) were demultiplexed using the atlas-utils demultiplex subcommand in Biostack suits (version 0.0.1, https://github.com/jameslz/biostack-suits/releases) with the barcode sequences. The pair-end reads were merged using the USEARCH fastq_mergepairs command (Edgar and Flyvbjerg, 2015) and subsequently trimmed. Operational taxonomic units (OTUs) were clustered at a 97% identity level using USEARCH UPARSE. The UPARSE pipeline analysis (Edgar, 2013) was used to discard chimeric sequences. The phylogenetic affiliation of 16S rRNA gene sequences was determined using the USEARCH SINTAX algorithm with the RDP training set (Cole et al., 2014; Edgar, 2016). OTUs annotated as mitochondrial or chloroplast rRNA gene fragments were excluded.
Microbial diversity indices, such as Shannon and Simpson, of sediment samples were analyzed using the USEARCH alpha_div (Edgar, 2010). Principal co-ordinates analysis (PCoA) and redundancy analysis (RDA) were performed using the Canoco 5 software to identify the microbial community structure and its relationship with substrate availability. Cluster analysis, Anosim test, and Pearson correlation were conducted in the R environment. Network analyses were invoked to explore microbial co-occurrence. The program Gephi 0.9.2 was used for visualization. OTUs from four seasons with a relative abundance larger than 0.01% were included. The standard for key OTU in the co-occurring relationship analysis was Spearman’s ρ > 0.8 and p < 0.001, for both positive and negative correlations (Jiao et al., 2016). The keystone taxa in the network (strong interactions with other OTUs) were identified following Banerjee et al. (2018).
3.
Results
3.1
Sediment physical and chemical properties
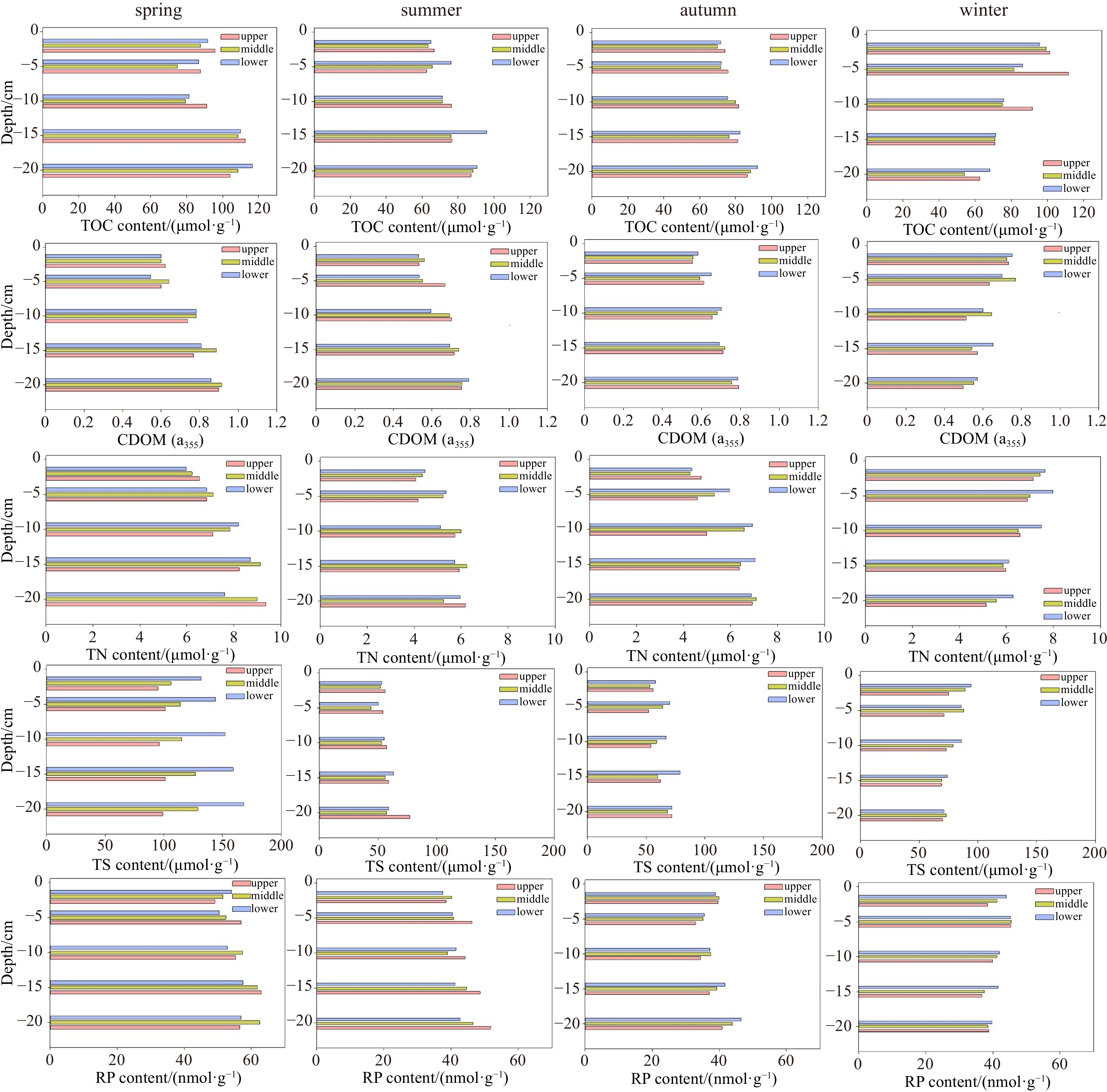
During spring, the average grain size of the sampled sediment ranged from 184 μm to 226 μm, showing a dominant proportion of sandy particles (>89%) in the sampled seepage face (Table S1). A minor variation in grain size was observed along the 20 cm depth seepage face and among seasons. TOC content in the sediment ranged from 76 μmol/g to 112 μmol/g during spring, with a peak at 15−20 cm depth (Fig. 2). Together with high TOC concentrations, CDOM (a355), TN and TS were also peaking at 15−20 cm depth during spring. During summer, the TOC concentration significantly decreased, dropping to the minimum of 62.5 μmol/g (Fig. 2). Concurrently, the concentration of remaining elements declined. During autumn and winter, TOC, TN, and TS concentrations increased, but remained lower than those obtained in the spring. The spatial and vertical variability of reactive phosphorus (RP) was minor among stations and different seasons.
Figure
2.
Profiles of total organic carbon (TOC), colored dissolved organic matter (CDOM), total nitrogen (TN), total sulfur (TS) and reactive phosphorus (RP) in the sediments (20 cm depth) of three tidal stations. CDOM concentrations in the tested water samples were estimated based on the absorption coefficient at 355 nm (Zhang et al., 2021).
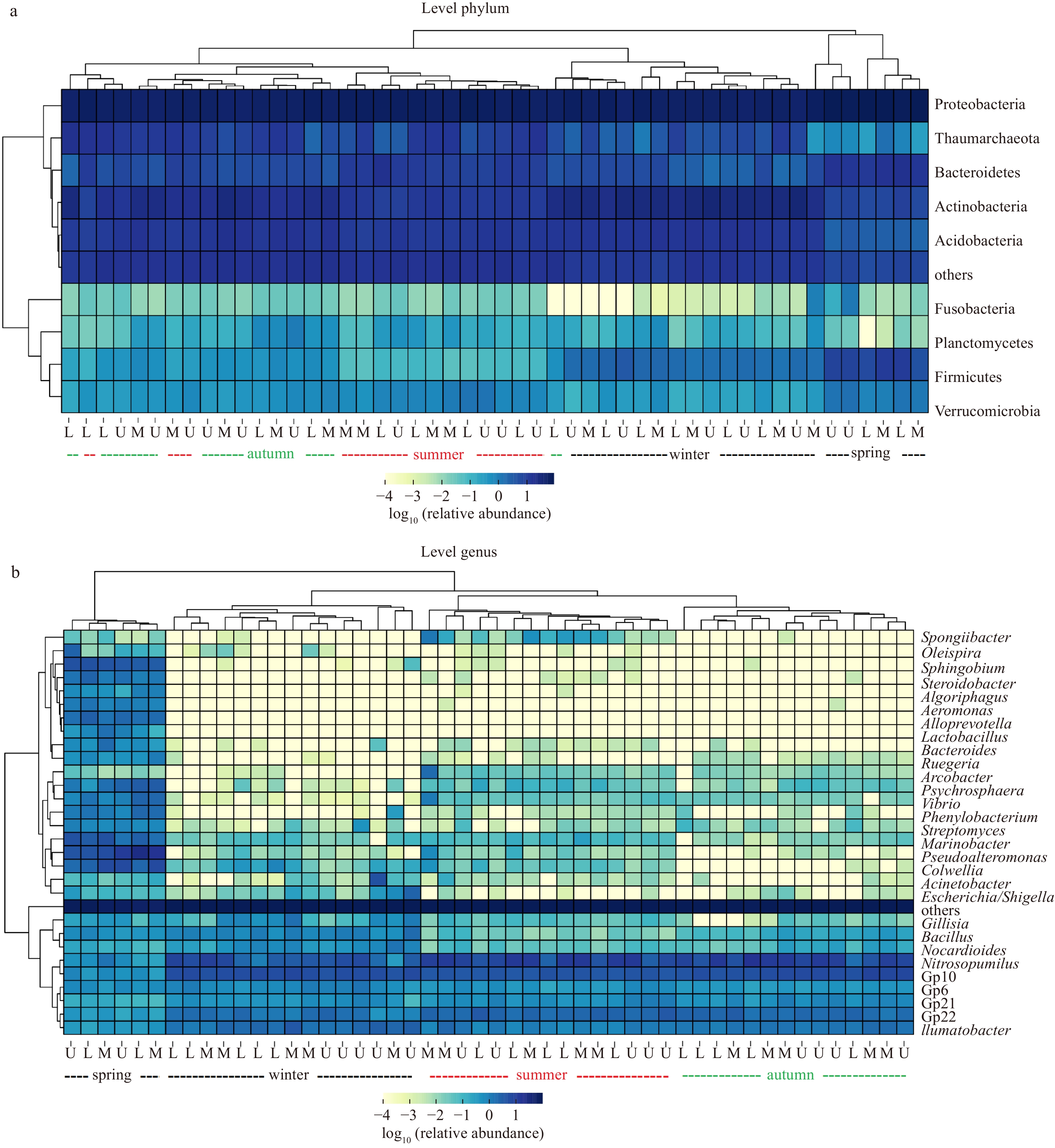
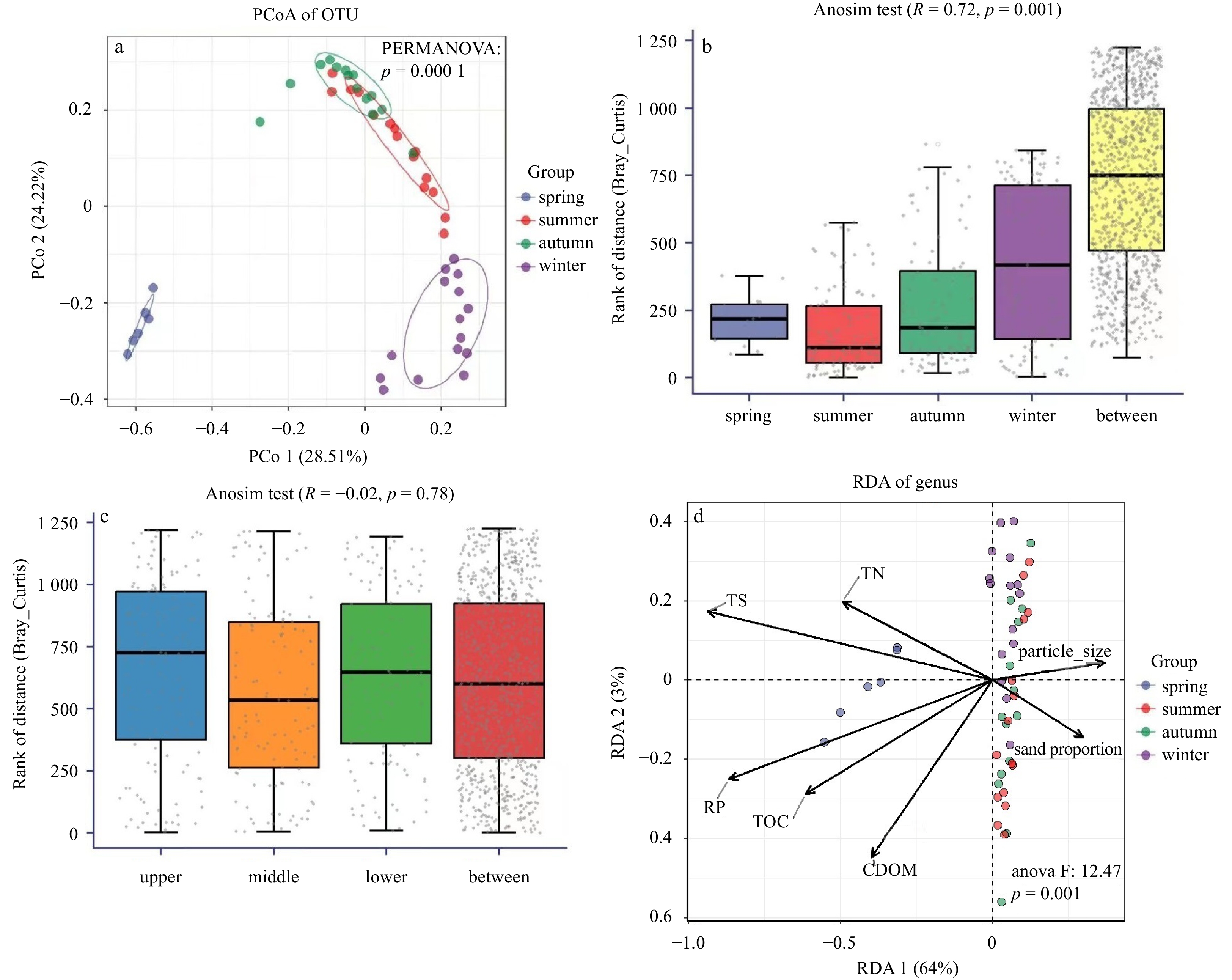
During spring, the OTUs identified in the sediment samples ranged from 1 288 to 1 411 (Table S2). In the summer, the quantity of sediment OTUs increased significantly, reaching 4 802. This increase in sediment OTUs during the summer season corresponded with higher values in the diversity indices, e.g., Shannon index (8.89 to 9.51) and Dominance index (0.988 to 0.994), as shown in Table S2. During the autumn and winter, the quantity of sediment OTUs varied from 934 to 6 060, respectively (Table S2), with the lowest value observed in the surface sediment during the winter despite TOC peaked in surface sediments in winter. The Shannon index ranged from 8.29 to 9.42 and 8.03 to 9.58 during these two seasons, while the Dominance index varied from 0.988 to 0.996. Among the sediment microbiota, the dominant phylum during all four seasons was Proteobacteria (Fig. 3a). Other notable phyla included Acidobacteria, Actinobacteria, and Bacteroidetes. At the genus level, the dominant genus was Pseudoalteromonas, while Gp10, Gp22, Bacillus, and Ilumatobacter were also enriched in the seepage face, as shown in Fig. 3b. Variations in relative abundance during the four seasons were observed, and different groups were identified via cluster analysis (Fig. 3) and PCoA (Fig. 4a). Compared to the significantly seasonal differences (p < 0.01; Fig. 4b), insignificant differences in microbial diversity were observed among the upper, middle, and lower intertidal stations at the OTU level (Fig. 4c).
Figure
3.
Biodiversity of benthic microorganism in the sampled seepage face at both phylum (top 10) and genus level (top 30). The darkness of the color indicates the relative abundance of each phylum or genus. Both figures separate microorganisms into four seasons, while microorganisms in different tidal boundaries (U: upper; M: middle; L: lower) are clearly mixed in the cluster analyses.
Figure
4.
Analyses of benthic microorganisms using different strategies. a. Principal co-ordinates analysis (PCoA) analysis for benthic microorganisms at operational taxonomic units (OTUs) level, which shows the significantly seasonal difference; b and c. results from Anosim tests to identify the difference between seasons and boundaries; d. the correlation between environmental factors and microorganisms at the genus level. RDA: redundancy analysis; TOC: total organic carbon; CDOM: colored dissolved organic matter; TN: total nitrogen; TS: total sulfur; RP: reactive phosphorus.
3.3
Relationships between macro elements and microbiota
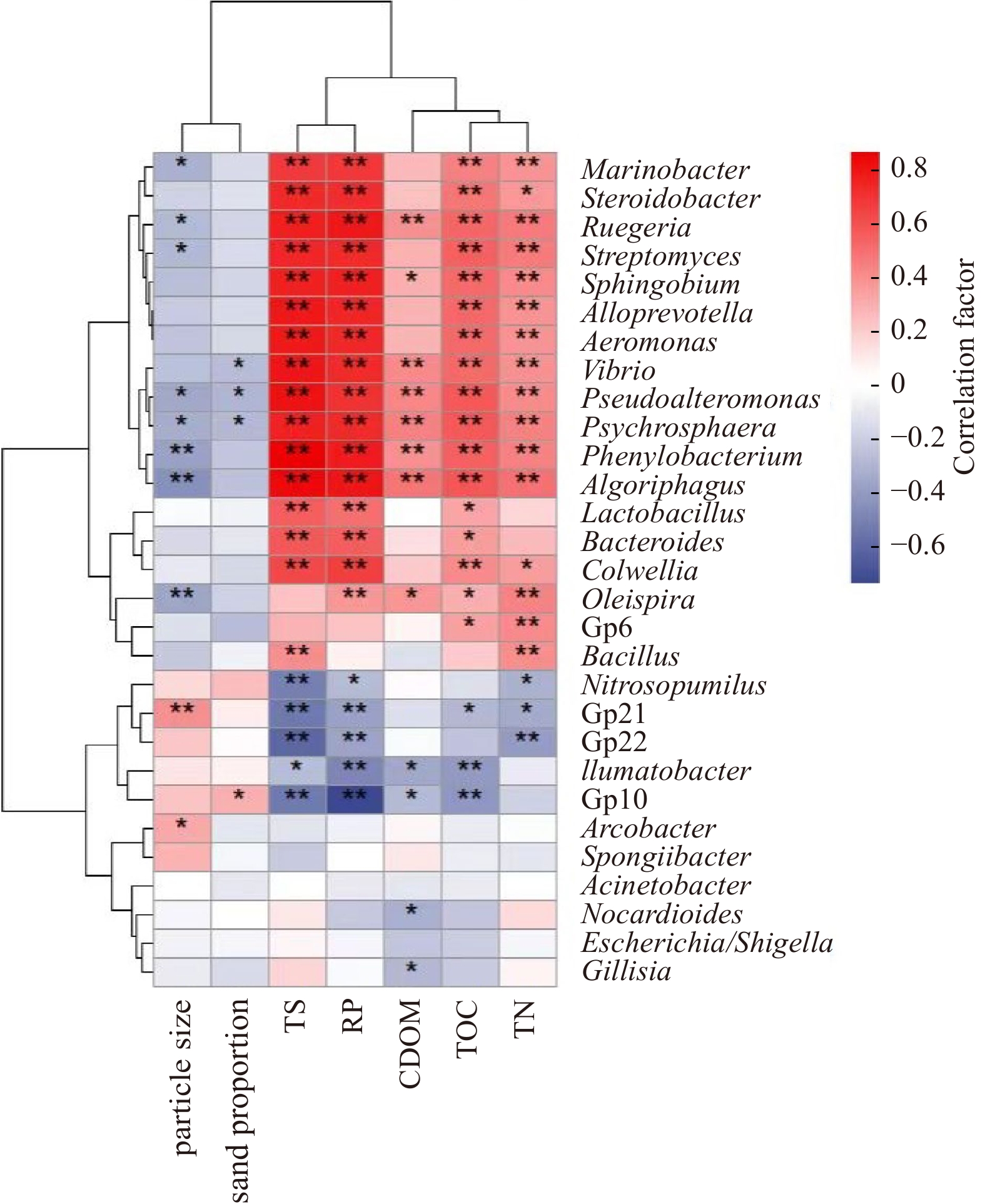
The present analysis aimed to investigate the relationship between sediment microbiota at the genus level and elemental substrates (TOC, TN, TS, and RP) in STE sediments across the sampled four seasons. RDA analysis (Fig. 4d) was implemented to outline this relationship. The findings revealed that TOC content significantly influenced the distribution of microbiota at the genus level (p < 0.01), particularly during the spring due to TOC enrichment (Fig. 2). Pearson correlation analysis (Fig. 5) revealed significant correlations between TOC and a wide range of benthic microbiota at the genus level. Additionally, TN and TS contents were identified as significant influencing factors on the microbial distribution, including the genera Marinobacter, Steroidobacter, and Ruegeria. The strong connection between macro elements and benthic microbiota highlights the active metabolism occurring in the sampled STE. Among all these functions, enzymes associated with carbon metabolism accounted for >5% relative abundance (Fig. 6), indicating the importance of carbon in the benthic transformations. The biosynthetic pathways for amino acids and purine, related with nitrogen and phosphorus, respectively, were emerged as key functions during summer and autumn.
Figure
5.
Pearson correlation for microorganisms and different environment factors. Asterisk indicates significant (*, p < 0.05) and extreme significant (**, p < 0.01) differences. TN: total nitrogen; TOC: total organic carbon; CDOM: colored dissolved organic matter; RP: reactive phosphorus; TS: total sulfur.
In spring, a total of 36 OTUs of sediment microorganisms with a relative abundance greater than 0.01% were observed in the network. This resulted in a weak correlation between clusters (Fig. 7a), as evidenced by the low values of edges and degree-centralization (Table 1). In contrast, during the summer season, a substantial increase in the quantity of microbial OTUs was observed, which significantly enhanced the connection between benthic microorganisms. This was reflected in the significant increase in average-degree and degree-centralization (Table 1), indicating a strong connection in the benthic metabolism. The keystone taxa identified during this season were Proteobacteria, Thaumarchaeota, and Actinobacteria (Fig. 7b), which were mainly responsible for organic matter degradation and nitrification (Table S3). During autumn, the correlation among sediment microorganisms remained strong, although the quantity of OTUs in the network decreased. The degree-centralization and clustering coefficient decreased (Table 1). Benthic metabolism was concentrated within a few keystone taxa of Proteobacteria, Acidobacteriota, and Actinobacteria. During winter, the network tended to be loosely again and the quantity of keystone OTUs significantly dropped (Fig. 7d).
Figure
7.
Network analyses for benthic microorganisms among four seasons. The number on each cycle indicates the operational taxonomic unit (OTU) number and color indicates the different phyla. The size of each cycle is positively correlated with the quantity of connection with other OTUs.
STE plays a crucial role in regulating the solute composition of porewater and supports a diverse range of benthic microbiota with various metabolic functions (Fig. 6). However, our understanding of microbiota in STE environments is limited compared to estuaries and coastal oceans, on both publication quantities and site coverage. In this study, the dominant phylum observed in the Sanggou Bay was Proteobacteria, which is consistent with other STEs such as the Shengsi Island, China (Chen et al., 2020), the Gloucester Beach, USA (Hong et al., 2019) and the Awur Bay, Indonesia (Adyasari et al., 2019; Table 2). The proportion of Proteobacteria in these environments ranged from 35% to 50%, indicating their high adaptability to marine environments (high salinity and low-level soluble substrates). However, the relative abundance of opportunistic microorganisms varied significantly among global sites (Table 2). At the genus level, the variability of sediment microbiota was more pronounced across different sites globally. In the Sanggou Bay, the key genera were Pseudoalteromonas, Colwellia, and Sphingobium, which were not observed in other STEs located in the temperate zone. This suggests that microbiota in STE environments is sensitive to ambient environments, despite similarities in chemical settings such as pelagic organic matter deposition, periodical enrichment of nutrient-enriched groundwater, and high salinity (>30 in all seasons; Jiang et al., 2020). Apart from water temperature influences (from ca. 4℃ to 26℃ from winter to summer; Jiang et al., 2020), this sensitivity indicates that benthic microbiota may exhibit selectivity towards specific environmental parameters within a range, such as the organic carbon composition (sugar and amino acids) and TOC∶TN content ratio in the sediment.
Table
2.
A global comparison of microbial compositions in subterranean estuaries or sandy sediments in coastal zones
Note: The dominant phylum in Boehm et al. (2014) could not be identified in the present results. OTUs: operational taxonomic units. − represents no data.
4.2
Spatial variability and relationship with macro elements
In comparison to the significant differences in microbial diversity observed among distinct STE sites (Table 2), the spatial variability of microbiota along the vertical profile in the seepage face of Sanggou Bay STE was limited. Specifically, at the phylum level, Proteobacteria was the dominant group and its relative abundance varied by less than 3% among layers with different depths (Fig. 3a). Furthermore, the key genera were found to be identical along the sediment profile (Fig. 3b). The sediment microbiota at the genus level from different depths was attributed to the same cluster during each season, as evidenced by the Bray-Curtis distance analysis (Fig. S1) and PCoA analysis (Fig. 4a). Therefore, depth in the seepage face (0−20 cm) was not a key factor in the distribution of benthic microbiota in the sampled seepage face. This hypothesis was further supported by RDA at the genus level (Fig. 4a). These results contrast with the spatial distribution of microbiota observed in the Ría de Vigo, Spain (Calvo-Martin et al., 2022) and the Gloucester Point, USA (Hong et al., 2019), where significant differences were observed along the vertical profile. The similarity in biodiversity observed in our site is likely due to the homogeneity of environmental factors in the sampled seepage face, which is shallow in depth. As the outer regime of STE, the seepage face is highly influenced by tidal setup and wave pumping effects (Fig. 1), which deliver substantial pelagic materials to the highly permeable sediments and improve the mixing between these elements and sandy particles (Calvo-Martin et al., 2021). Consequently, the microbiota attached to the particle surface tends to be evenly distributed along the vertical profile.
The sediment parameters, particularly the organic carbon content, varied among different tidal boundaries (Fig. 2). The higher boundary of the seepage face, which frequently serves as the entry point of seawater (Robinson et al., 2007), received a significant amount of pelagic organic matter (Waska et al., 2021). Consequently, TOC and TN contents in the higher intertidal stations were more enriched compared to the median and lower stations, which could promote the growth of organic carbon degradation-related microorganisms, such as OTU-249 and OTU-211 (Table S3). In comparison, only slight differences in the relative abundance of microorganisms were observed among boundaries in the sampled seepage face at the phylum level (Fig. 3a), likely due to the versatile metabolic pathways. The diversity variation was pronounced at the genus level, particularly for Pseudoalteromonas and Colwellia, species that preferring organic carbon enrichment. Nevertheless, according to the Anosim test, the difference in microbial community composition among boundaries was still insignificant (Fig. 4c). This distribution pattern of sediment microorganisms appears to contradict the substrate preference of microorganisms. As a region regularly flushed by seawater, porewater intensively transports microorganisms and their spores (spore-forming bacteria mainly to be Bacillus and Clostridium) among different boundaries (Lee et al., 2017). This hydraulic cycling facilitates the transport of microbial species, thereby enhancing the homogeneity of biodiversity. Additionally, the microorganisms obtained from the seepage face may also rely on the dissolved fraction of organic carbon and nutrients. In various in situ observations, the addition of labile organic matter, such as glucose and amino acids, to coastal sediments significantly enhanced microbial diversity (Jessen et al., 2017) and activity (Jiang et al., 2016). Compared with sediment fractions, dissolved solute contents in porewater can be freely transported under the effect of tidal pumping and utilized by the microbiota among different boundaries. Taken together, the weak spatial variability among boundaries in the studied seepage face was observed.
4.3
Seasonal variability and drivers
Compared to the insignificance observed among boundaries, a significant variation was detected among four seasons using the Anoism test (Fig. 4b). Furthermore, in the PCoA (Fig. 4a) and RDA (Fig. 4d), benthic microorganisms were distinctly separated into four clusters, indicating a strong seasonality. In estuaries or subterranean estuaries, the benthic microbial diversity, even at the phylum level, frequently varied drastically among different months. For example, in the Rajang Estuary, Malaysia, the dominant phyla during August included Proteobacteria, Firmicutes, Bacteroidetes and Deinococcus-Thermus, while the relative abundance of Deinococcus-Thermus in freshwater endmember markedly decreased in September compared with the records found in March and August (Sia et al., 2019). Similar variations in the relative abundance of Bacteroidetes between seasons were also observed in the Mandovi Estuary, India (Fernandes et al., 2022). In the Ría de Vigo STE (Spain), the dominant phylum changed from Proteobacteria to Nanoarchaeaeota between March and July (Calvo-Martin et al., 2022). In the Sanggou Bay STE, the dominant phylum remained Proteobacteria during all four seasons, while its relative abundance significantly varied among seasons (Fig. 3a). At the genus level, the seasonal change tended to be more drastic. The dominant genus significantly varied between spring and the other three seasons (Fig. 3b). For such seasonality, temperature would be the key factor modulating the microbial diversity since the preferential temperature range varied among microbiota species. For instance, the labile organic carbon degradation-related microorganisms in the Umbrella Creek, USA, preferred the ambient temperature from 22℃ to 34℃ (Weston and Joye, 2005). In the Sanggou Bay, the porewater temperature was constant in the seepage face during each season, varying from 27.6℃ in summer to 4.2℃ during winter (Jiang et al., 2020). Such large changes could lead to the change of the growth rate of different sediment microbiota. At the OTU level (Table S3), several keystone taxa obtained from the network analyses, such as Tetrasphaera australiensis (OTU-249) and Pelagibius litoralis (OTU-160), were identified as temperature-sensitive (Liu et al., 2022; Choi et al., 2009), which likely led to the seasonal variations observed in the relative abundance. However, seawater and environmental temperatures between spring and autumn in our sampling site were similar, around 20℃ (Jiang et al., 2020). Additionally, the porewater exchange rate and salinity in different boundaries were also similar between these two seasons (Jiang et al., 2023), while the microbiota diversity was markedly different from phylum to OTU level between these two seasons, revealing the significance from the substrate supply on shaping the microbial community structure.
As aforementioned, oceanic compounds, including macroalgae and plankton debris, are crucial sources of material for seepage faces (Jiang et al., 2020). The concentrations of TOC, TN and TS in the seepage face during autumn only accounted for 40% to 60% of the records during spring, compared to the substrate concentration variations in different depths and boundaries. Furthermore, substrate variability from different seasons has a long-term impact on benthic microbiota in coastal sediments (Wei et al., 2022), covering several spring-neap tidal cycles, which significantly shapes the community structure. During spring, the bay water was enriched with pelagic organic matter due to the phytoplankton bloom, evidenced by high levels of particulate organic carbon (POC) and chlorophyll a in coastal water during high tides (Jiang et al., 2020). Additionally, kelp harvest was frequently conducted during late spring (Zeng et al., 2015), and degradation of kelp debris added carbon matter content in the bay water, especially labile organic matter such as amino acids and oligosaccharides (Clark et al., 2016). In response to this scenario, heterotrophic microorganisms, such as Sphingobium, Pseudoalteromonas, and Colwellia, were significantly enriched in the seepage face. However, during spring, the benthic metabolism was loosely connected via few OTUs (Fig. 7a), indicating the absence of keystone OTUs. This weak connection for benthic microorganisms was also observed in an incubation experiment using soils from the Huanghe River delta, where organic matter in soils was significantly enhanced via adding reed straw (Yu et al., 2022). This distribution pattern revealed that the benthic microbiota tended to be independent and freely metabolizing during spring by selecting suitable substrates from the rich substrate “pool”. In contrast, during autumn, the carbon supply from oceanic sources substantially shrank and decrease TOC content in sediments (Fig. 2). The benthic microbiota in the seepage face showed a rapid response to such changes. The microbial network was concentrated around key OTUs, and a significant quantity of strong correlations was obtained, indicating that the benthic metabolism during autumn displayed coordination (positive linkage) and competition (negative linkage) between species (cf. low carbon sediment in the Changjiang River Estuary; Wei et al., 2022). Moreover, at the genus level, a significant decrease in a wide range of heterotrophic microorganisms, such as Phenylobacterium, Colwellia, Gillisia, and Acinetobacter, was obtained. In contrast, the relative abundance of autotrophic species, such as Nitrosopumilus and Thioprofundum, increased (Fig. 3b), indicating that the species with the function of nitrogen and sulfur oxidation also became key players to sustain the benthic metabolism function.
In the summer season, characterized by high temperatures and abundant pelagic organic matter, a distinct pattern was observed in comparison to both spring and autumn. A significant quantity of microbial OTUs was obtained (Table S2), and the number of keystone taxa in the metabolism network markedly increased. Proteobacteria, Actinobacteria, and Thaumarchaeota played a crucial role in biogeochemical transformations (Fig. 7b), primarily through organic matter degradation (identified as carbon metabolism in Fig. 6). In addition, the metabolism related with N and P was also active. During summer, the connection between these species and ambient microorganisms was significantly enhanced, resulting in the densest network (Fig. 7b). Phytoplankton concentrations remained high during summer, while dinoflagellates biomass in bay water increased (Yuan et al., 2014). Compared to diatoms, dinoflagellates included several species that produced biological toxins (Liu et al., 2023), making their degradation in benthic environments more complex. High temperatures accelerated the growth of benthic microbiota and enhanced competition in heterotrophic degradation, leading to clustering in the metabolism network and fitting into different niches. The strong competition in organic matter degradation also benefited autotrophic microorganisms in acquiring nitrogen and sulfur compounds. As a response, the relative abundance of autotrophic microorganisms, especially nitrifiers, peaked in summer and acted as keystone taxa in the OTU network (Table S3). During winter, the low temperature depressed the growth and activity of benthic microorganisms, leading to the accumulation of organic matter in sandy sediments via slowing degradation rate. The microbial network tended to be loosely connected, and the number of keystone OTUs sharply decreased.
5.
Conclusions
This study investigated the microbiota within the sediment in the Sanggou Bay STE across four seasons, as benthic microorganisms acted as a modulator for solutes in porewater and attached materials on sediment particles. Proteobacteria was found to be the dominant phylum throughout all seasons, while opportunistic phyla Actinobacteria, Acidobacteria, Bacteroidetes, and Thaumarchaeota varied. Heterotrophic microbiota constituted the majority of these microorganisms, indicating that the degradation of pelagic organic matter served as the primary energy source for benthic metabolism. Notably, the substrate condition, rather than environmental temperature was found to significantly shape the benthic microbial network in the sampled seepage face, especially via the comparison between spring and autumn. During spring and winter, benthic substrate supply, particularly organic matter, surpassed the quantity of benthic microorganisms, resulting in loosely connection with independent metabolism. In contrast, during summer and autumn, the benthic metabolism network was concentrated into one or two clusters with tight correlations among keystone taxa. This pattern may indicate an enhanced utilization of pelagic organic matter via co-degradation and direct competition during substrate shortage and microbial enrichment. Furthermore, the relative deficiency of organic matter promoted the growth of autotrophic microorganisms involved in the oxidation of nitrogen and sulfur compounds. In summary, the balance between substrate supply, primarily pelagic compounds, and microbial biomass was crucial in shaping the benthic microbial network.
Acknowledgements:
We thank Guosen Zhang and Ying Cui in East China Normal University, and Guanghui Lin in Tsinghua University for laboratory analyses, and Lei Zhang from DeepBiome Co., Ltd. for bioinformatic assistance. We are thankful to the editor and two anonymous reviewers whose comments helped to improve an earlier version of the manuscript.
Adyasari D, Hassenrück C, Oehler T, et al. 2019. Microbial community structure associated with submarine groundwater discharge in northern Java (Indonesia). Science of the Total Environment, 689: 590–601. doi: 10.1016/j.scitotenv.2019.06.193
Archana A, Francis C A, Boehm A B. 2021. The beach aquifer microbiome: research gaps and data needs. Frontiers in Environmental Science, 9: 653568. doi: 10.3389/fenvs.2021.653568
Banerjee S, Schlaeppi K, van der Heijden M G A. 2018. Keystone taxa as drivers of microbiome structure and functioning. Nature Reviews Microbiology, 16(9): 567–576. doi: 10.1038/s41579-018-0024-1
Boehm A B, Yamahara K M, Sassoubre L M. 2014. Diversity and transport of microorganisms in intertidal sands of the California coast. Applied and Environmental Microbiology, 80(13): 3943–3951
Bohórquez J, McGenity T J, Papaspyrou S, et al. 2017. Different types of diatom-derived extracellular polymeric substances drive changes in heterotrophic bacterial communities from intertidal sediments. Frontiers in microbiology, 8: 245
Calvo-Martin E, Álvarez-Salgado X A, Rocha C, et al. 2021. Reactive solute transport through two contrasting subterranean estuary exit sites in the Ría de Vigo (NW Iberian Peninsula). Frontiers in Marine Science, 8: 626813. doi: 10.3389/fmars.2021.626813
Calvo-Martin E, Teira E, Álvarez-Salgado X A, et al. 2022. On the hidden diversity and niche specialization of the microbial realm of subterranean estuaries. Environmental Microbiology, 24(12): 5859–5881. doi: 10.1111/1462-2920.16160
Chen Xiaogang, Ye Qi, Sanders C J, et al. 2020. Bacterial-derived nutrient and carbon source-sink behaviors in a sandy beach subterranean estuary. Marine Pollution Bulletin, 160: 111570. doi: 10.1016/j.marpolbul.2020.111570
Cho H M, Kim G, Kwon E Y, et al. 2018. Radium tracing nutrient inputs through submarine groundwater discharge in the global ocean. Scientific Reports, 8(1): 2439. doi: 10.1038/s41598-018-20806-2
Choi D H, Hwang C Y, Cho B C. 2009. Pelagibius litoralis gen. nov., sp. nov., a marine bacterium in the family Rhodospirillaceae isolated from coastal seawater. International Journal of Systematic and Evolutionary Microbiology, 59(4): 818–823. doi: 10.1099/ijs.0.002774-0
Clark C D, De Bruyn W J, Aiona P D. 2016. Temporal variation in optical properties of chromophoric dissolved organic matter (CDOM) in Southern California coastal waters with nearshore kelp and seagrass. Limnology and Oceanography, 61(1): 32–46. doi: 10.1002/lno.10198
Cole J R, Wang Qiong, Fish J A, et al. 2014. Ribosomal Database Project: data and tools for high throughput rRNA analysis. Nucleic Acids Research, 42(D1): D633–D642. doi: 10.1093/nar/gkt1244
Edgar R C. 2010. Search and clustering orders of magnitude faster than BLAST. Bioinformatics, 26(19): 2460–2461. doi: 10.1093/bioinformatics/btq461
Edgar R C. 2013. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nature Methods, 10(10): 996–998. doi: 10.1038/nmeth.2604
Edgar R C, Flyvbjerg H. 2015. Error filtering, pair assembly and error correction for next-generation sequencing reads. Bioinformatics, 31(21): 3476–3482. doi: 10.1093/bioinformatics/btv401
Fernandes S O, Gonsalves M J, Nazareth D R, et al. 2022. Seasonal variability in environmental parameters influence bacterial communities in mangrove sediments along an estuarine gradient. Estuarine, Coastal and Shelf Science, 270: 107791
Hong Yiguo, Wu Jiapeng, Wilson S, et al. 2019. Vertical stratification of sediment microbial communities along geochemical gradients of a subterranean estuary located at the Gloucester Beach of Virginia, United States. Frontiers in Microbiology, 9: 3343. doi: 10.3389/fmicb.2018.03343
Jessen G L, Lichtschlag A, Ramette A, et al. 2017. Hypoxia causes preservation of labile organic matter and changes seafloor microbial community composition (Black Sea). Science Advances, 3(2): e1601897. doi: 10.1126/sciadv.1601897
Jiang Shan, Jin Jie, Wei Yongjun, et al. 2023. Sandy seepage faces as bioactive nitrate reactors: Biogeochemistry, microbial ecology and metagenomics. Geoscience Frontiers, 14(3): 101529. doi: 10.1016/j.gsf.2022.101529
Jiang Shan, Jin Jie, Wu Ying, et al. 2021a. Response of Nitrate Processing to Bio-labile Dissolved Organic Matter Supply Under Variable Oxygen Conditions in a Sandy Beach Seepage Face. Frontiers in Marine Science, 8: 642143. doi: 10.3389/fmars.2021.642143
Jiang Shan, Kavanagh M, Ibánhez J S P, et al. 2021b. Denitrification-nitrification process in permeable coastal sediments: An investigation on the effect of salinity and nitrate availability using flow-through reactors. Acta Oceanologica Sinica, 40(9): 1–12. doi: 10.1007/s13131-021-1811-5
Jiang Shan, Lu Haoliang, Zhang Qiong, et al. 2016. Effect of enhanced reactive nitrogen availability on plant-sediment mediated degradation of polycyclic aromatic hydrocarbons in contaminated mangrove sediment. Marine Pollution Bulletin, 103(1–2): 151–158
Jiang Shan, Zhang Yixue, Jin Jie, et al. 2020. Organic carbon in a seepage face of a subterranean estuary: turnover and microbial interrelations. Science of the Total Environment, 725: 138220. doi: 10.1016/j.scitotenv.2020.138220
Jiao Shuo, Liu Zhenshan, Lin Yanbing, et al. 2016. Bacterial communities in oil contaminated soils: biogeography and co-occurrence patterns. Soil Biology and Biochemistry, 98: 64–73. doi: 10.1016/j.soilbio.2016.04.005
Klier J, Dellwig O, Leipe T, et al. 2018. Benthic bacterial community composition in the oligohaline-marine transition of surface sediments in the Baltic Sea based on rRNA analysis. Frontiers in Microbiology, 9: 236
Lee E, Shin D, Hyun S P, et al. 2017. Periodic change in coastal microbial community structure associated with submarine groundwater discharge and tidal fluctuation. Limnology and Oceanography, 62(2): 437–451. doi: 10.1002/lno.10433
Liang Jiawei, Mai Wenning, Tang Jinfeng, et al. 2019. Highly effective treatment of petrochemical wastewater by a super-sized industrial scale plant with expanded granular sludge bed bioreactor and aerobic activated sludge. Chemical Engineering Journal, 360: 15–23. doi: 10.1016/j.cej.2018.11.167
Liang Jiawei, Mai Wenning, Wang Jia, et al. 2021. Performance and microbial communities of a novel integrated industrial-scale pulp and paper wastewater treatment plant. Journal of Cleaner Production, 278: 123896. doi: 10.1016/j.jclepro.2020.123896
Linkhorst A, Dittmar T, Waska H. 2017. Molecular fractionation of dissolved organic matter in a shallow subterranean estuary: the role of the iron curtain. Environmental Science & Technology, 51(3): 1312–1320
Liu Xiaohan, Liu Yuyang, Chai Zhaoyang, et al. 2023. A combined approach detected novel species diversity and distribution of dinoflagellate cysts in the Yellow Sea, China. Marine Pollution Bulletin, 187: 114567. doi: 10.1016/j.marpolbul.2022.114567
Liu Hongjun, Zeng Wei, Meng Qingan, et al. 2022. Phosphorus removal performance, intracellular metabolites and clade-level community structure of Tetrasphaera-dominated polyphosphate accumulating organisms at different temperatures. Science of the Total Environment, 842: 156913. doi: 10.1016/j.scitotenv.2022.156913
Moore W S. 2010. The effect of submarine groundwater discharge on the ocean. Annual Review of Marine Science, 2: 59–88. doi: 10.1146/annurev-marine-120308-081019
Reckhardt A, Beck M, Seidel M, et al. 2015. Carbon, nutrient and trace metal cycling in sandy sediments: a comparison of high-energy beaches and backbarrier tidal flats. Estuarine, Coastal and Shelf Science, 159: 1–14
Robinson C, Gibbes B, Carey H, et al. 2007. Salt-freshwater dynamics in a subterranean estuary over a spring-neap tidal cycle. Journal of Geophysical Research: Oceans, 112(C9): C09007
Rocha C, Jiang Shan, Ibánhez J S P, et al. 2022. The effects of subterranean estuary dynamics on nutrient resource ratio availability to microphytobenthos in a coastal lagoon. Science of the Total Environment, 851: 157522. doi: 10.1016/j.scitotenv.2022.157522
Rocha C, Robinson C E, Santos I R, et al. 2021. A place for subterranean estuaries in the coastal zone. Estuarine, Coastal and Shelf Science, 250: 107167
Ruiz-González C, Rodríguez-Pie L, Maister O, et al. 2022. High spatial heterogeneity and low connectivity of bacterial communities along a Mediterranean subterranean estuary. Molecular Ecology, 31(22): 5745–5764. doi: 10.1111/mec.16695
Santos I R, Chen Xiaogang, Lecher A L, et al. 2021. Submarine groundwater discharge impacts on coastal nutrient biogeochemistry. Nature Reviews Earth & Environment, 2(5): 307–323
Sia E S A, Zhu Zhuoyi, Zhang Jing, et al. 2019. Biogeographical distribution of microbial communities along the Rajang River-South China Sea continuum. Biogeosciences, 16(21): 4243–4260. doi: 10.5194/bg-16-4243-2019
Wang Liping, Zheng Binghui, Nan Bingxu, et al. 2014. Diversity of bacterial community and detection of nirS-and nirK-encoding denitrifying bacteria in sandy intertidal sediments along Laizhou Bay of Bohai Sea, China. Marine Pollution Bulletin, 88(1−2): 215–223. doi: 10.1016/j.marpolbul.2014.09.002
Waska H, Simon H, Ahmerkamp S, et al. 2021. Molecular traits of dissolved organic matter in the subterranean estuary of a high-energy beach: indications of sources and sinks. Frontiers in Marine Science, 8: 607083. doi: 10.3389/fmars.2021.607083
Wei Yongjun, Jiang Shan, Tian Lingmin, et al. 2022. Benthic microbial biogeography along the continental shelf shaped by substrates from the Changjiang River plume. Acta Oceanologica Sinica, 41(1): 118–131. doi: 10.1007/s13131-021-1861-8
Wei Yongjun, Wu Yuqian, Zhang Lei, et al. 2020. Genome recovery and metatranscriptomic confirmation of functional acetate-oxidizing bacteria from enriched anaerobic biogas digesters. Environmental Pollution, 265: 114843. doi: 10.1016/j.envpol.2020.114843
Weston N B, Joye S B. 2005. Temperature-driven decoupling of key phases of organic matter degradation in marine sediments. Proceedings of the National Academy of Sciences of the United States of America, 102(47): 17036–17040
Wu Ying, Zhu Kun, Zhang Jing, et al. 2019. Distribution and degradation of terrestrial organic matter in the sediments of peat-draining rivers, Sarawak, Malaysian Borneo. Biogeosciences, 16(22): 4517–4533. doi: 10.5194/bg-16-4517-2019
Yu Lu, Bai Junhong, Huang Laibin, et al. 2022. Carbon-rich substrates altered microbial communities with indication of carbon metabolism functional shifting in a degraded salt marsh of the Yellow River Delta, China. Journal of Cleaner Production, 331: 129898. doi: 10.1016/j.jclepro.2021.129898
Yuan Mingli, Zhang Cuixia, Jiang Zengjie, et al. 2014. Seasonal variations in phytoplankton community structure in the Sanggou, Ailian, and Lidao Bays. Journal of Ocean University of China, 13(6): 1012–1024. doi: 10.1007/s11802-014-2305-2
Zeng Dingyong, Huang Daji, Qiao Xudong, et al. 2015. Effect of suspended kelp culture on water exchange as estimated by in situ current measurement in Sanggou Bay, China. Journal of Marine Systems, 149: 14–24. doi: 10.1016/j.jmarsys.2015.04.002
Zhang Yixue, Wu Ying, Zhang Jing, et al. 2021. Seasonal dynamics of surface dissolved organic matter in the South China Sea and the Straits of Malacca: Implications for biogeochemical province delineation of marginal seas. Journal of Geophysical Research: Biogeosciences, 126(10): e2021JG006535. doi: 10.1029/2021JG006535
Yueming Wu, Beiyun Xu, Danqing Sun, et al. Higher dynamic and turnover yet enhanced organic carbon burial in Spartina alterniflora sediments: A case study in a temperate coastal wetland. CATENA, 2025, 249: 108701. doi:10.1016/j.catena.2025.108701
Yueming Wu, Yongjun Wei, Juan Severino Pino Ibánhez, Danqing Sun, Kaiqi Lu, Qianqian Wang, Jian Li, Shan Jiang. Microbiota of a seepage face at the mouth of a subterranean estuary: diversity, distribution and substrate dependence[J]. Acta Oceanologica Sinica, 2023, 42(8): 147-157. doi: 10.1007/s13131-023-2257-8
Yueming Wu, Yongjun Wei, Juan Severino Pino Ibánhez, Danqing Sun, Kaiqi Lu, Qianqian Wang, Jian Li, Shan Jiang. Microbiota of a seepage face at the mouth of a subterranean estuary: diversity, distribution and substrate dependence[J]. Acta Oceanologica Sinica, 2023, 42(8): 147-157. doi: 10.1007/s13131-023-2257-8
Note: The dominant phylum in Boehm et al. (2014) could not be identified in the present results. OTUs: operational taxonomic units. − represents no data.
Note: The dominant phylum in Boehm et al. (2014) could not be identified in the present results. OTUs: operational taxonomic units. − represents no data.
Figure 1. Sketch of subterranean estuary (STE) and seepage face in coastal zones. This figure highlights the three sampling sites (upper, middle and lower tidal stations) and major organic matter (OM) sources for STE. This figure was modified from Jiang et al. (2021a).
Figure 2. Profiles of total organic carbon (TOC), colored dissolved organic matter (CDOM), total nitrogen (TN), total sulfur (TS) and reactive phosphorus (RP) in the sediments (20 cm depth) of three tidal stations. CDOM concentrations in the tested water samples were estimated based on the absorption coefficient at 355 nm (Zhang et al., 2021).
Figure 3. Biodiversity of benthic microorganism in the sampled seepage face at both phylum (top 10) and genus level (top 30). The darkness of the color indicates the relative abundance of each phylum or genus. Both figures separate microorganisms into four seasons, while microorganisms in different tidal boundaries (U: upper; M: middle; L: lower) are clearly mixed in the cluster analyses.
Figure 4. Analyses of benthic microorganisms using different strategies. a. Principal co-ordinates analysis (PCoA) analysis for benthic microorganisms at operational taxonomic units (OTUs) level, which shows the significantly seasonal difference; b and c. results from Anosim tests to identify the difference between seasons and boundaries; d. the correlation between environmental factors and microorganisms at the genus level. RDA: redundancy analysis; TOC: total organic carbon; CDOM: colored dissolved organic matter; TN: total nitrogen; TS: total sulfur; RP: reactive phosphorus.
Figure 5. Pearson correlation for microorganisms and different environment factors. Asterisk indicates significant (*, p < 0.05) and extreme significant (**, p < 0.01) differences. TN: total nitrogen; TOC: total organic carbon; CDOM: colored dissolved organic matter; RP: reactive phosphorus; TS: total sulfur.
Figure 6. Benthic microbial functions during summer and autumn, identified via metagenomic analysis. TCA: tricarboxylic acid cycle.
Figure 7. Network analyses for benthic microorganisms among four seasons. The number on each cycle indicates the operational taxonomic unit (OTU) number and color indicates the different phyla. The size of each cycle is positively correlated with the quantity of connection with other OTUs.




 DownLoad:
DownLoad:






 DownLoad:
DownLoad: