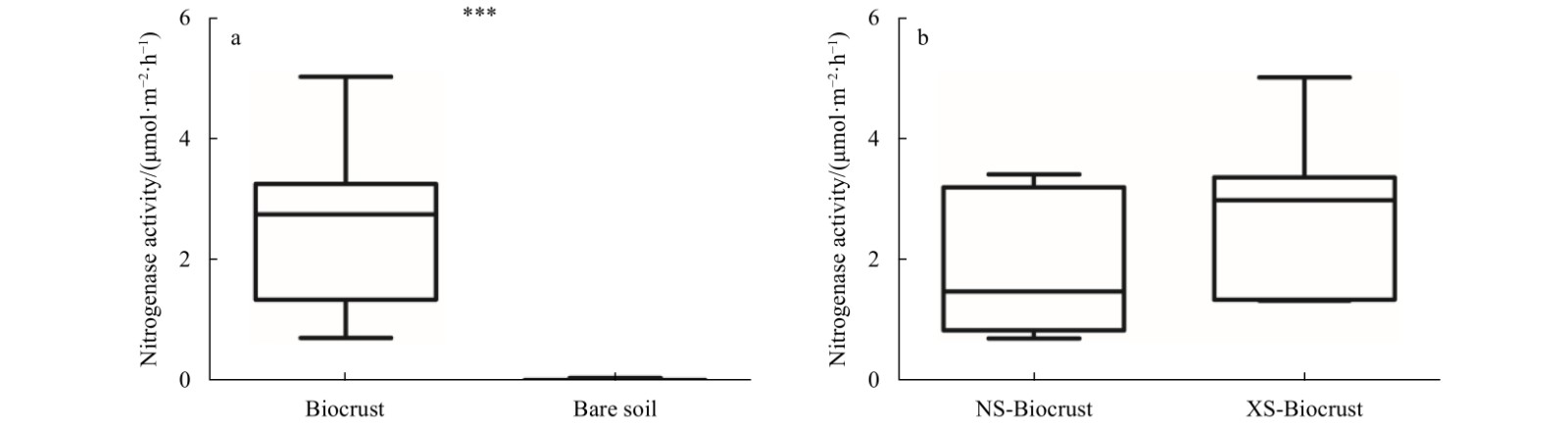
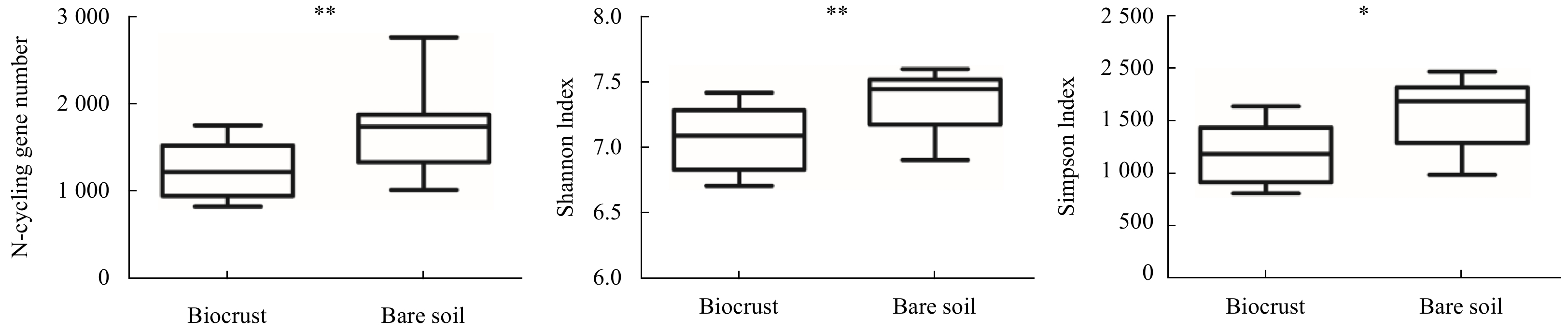
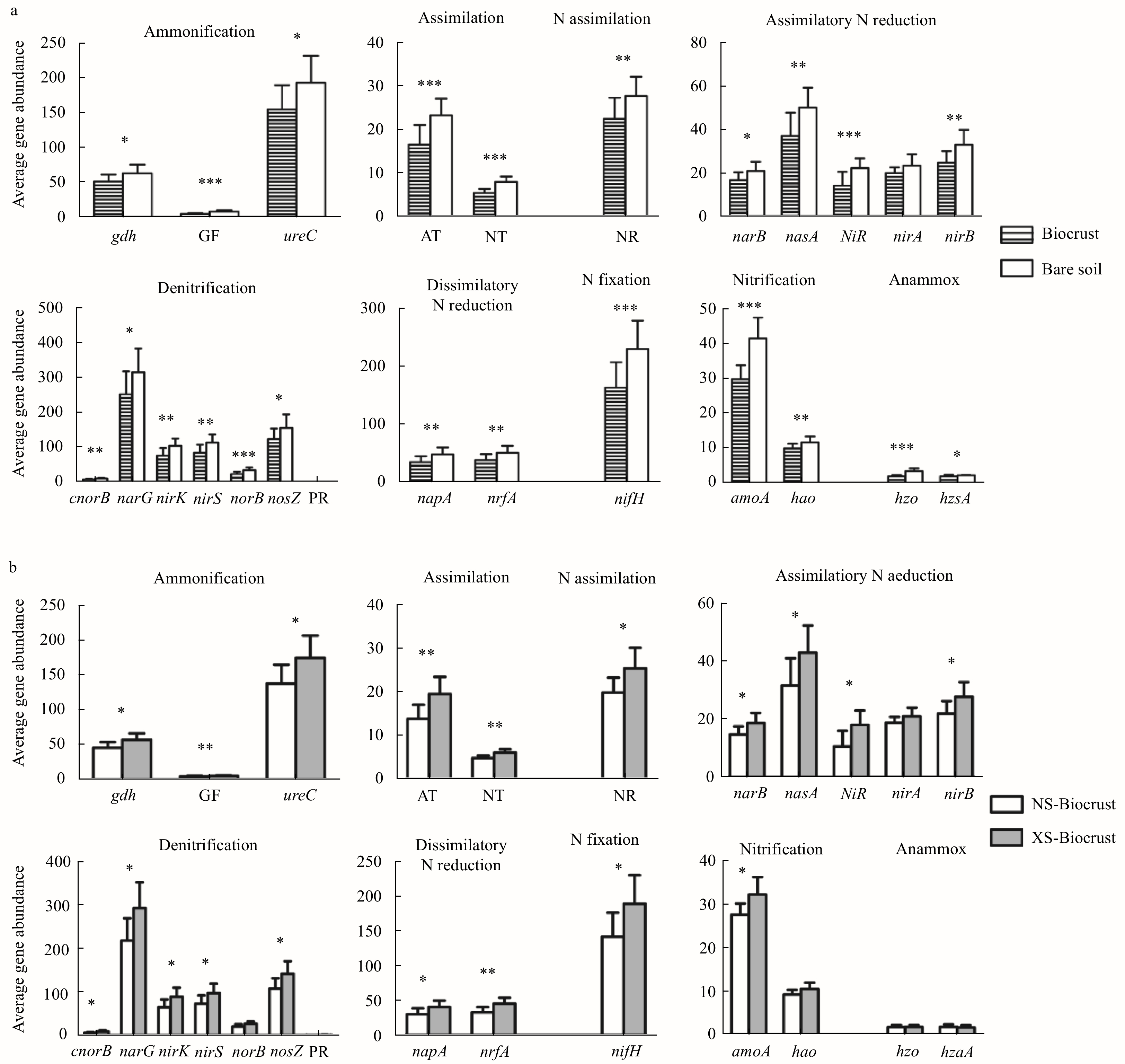
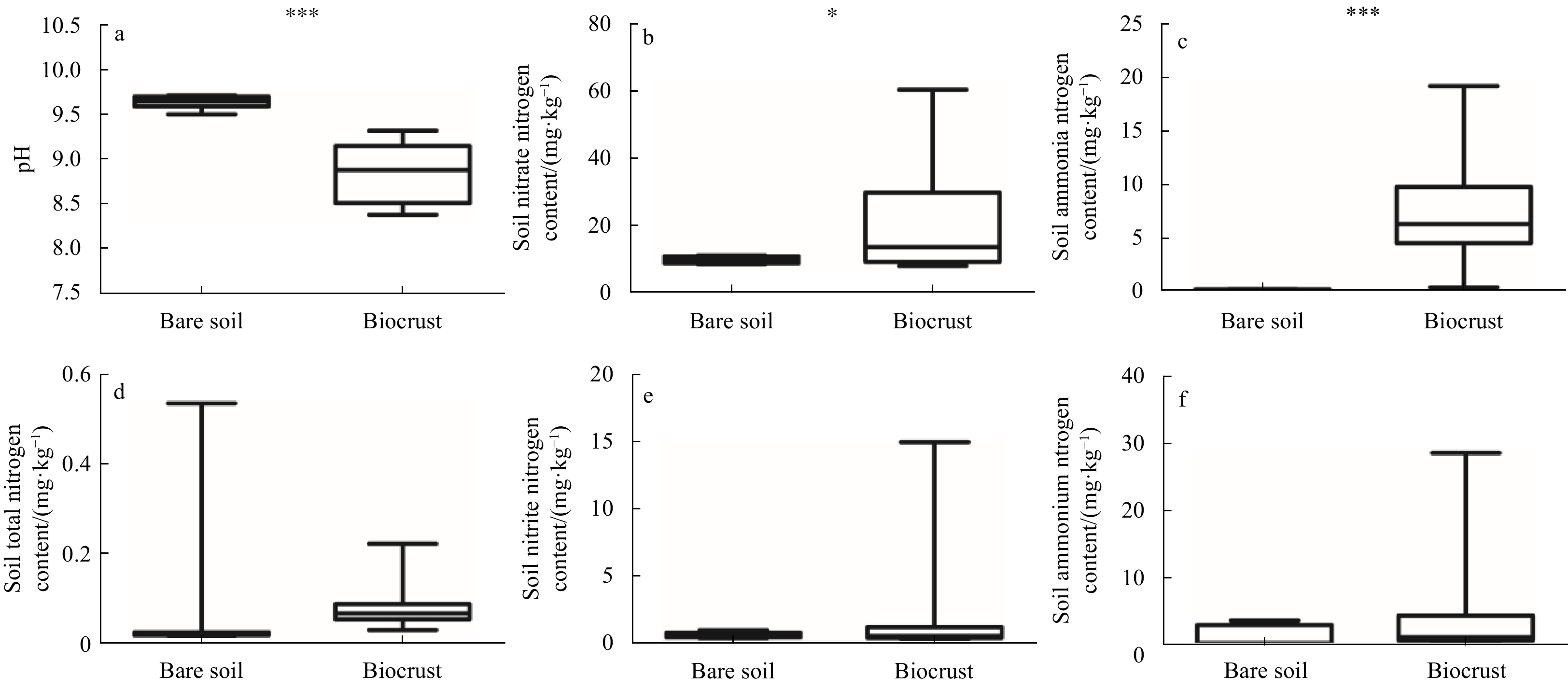
| Citation: | Lin Wang, Si Zhang, Jie Li. The role of biocrusts in nitrogen cycling on the tropical reef islands, South China Sea[J]. Acta Oceanologica Sinica, 2021, 40(4): 116-126. doi: 10.1007/s13131-021-1783-5

|

| [1] |
Acosta-Martínez V, Cruz L, Sotomayor-Ramírez D, et al. 2007. Enzyme activities as affected by soil properties and land use in a tropical watershed. Applied Soil Ecology, 35(1): 35–45. doi: 10.1016/j.apsoil.2006.05.012
|
| [2] |
Baran R, Brodie E L, Mayberry-Lewis J, et al. 2015. Exometabolite niche partitioning among sympatric soil bacteria. Nature Communications, 6: 8289. doi: 10.1038/ncomms9289
|
| [3] |
Barger N N, Belnap J, Ojima D S, et al. 2005. NO gas loss from biologically crusted soils in Canyonlands National Park, Utah. Biogeochemistry, 75(3): 373–391. doi: 10.1007/s10533-005-1378-9
|
| [4] |
Barger N N, Weber B, Garcia-Pichel F, et al. 2016. Patterns and controls on nitrogen cycling of biological soil crusts. In: Weber B, Büdel B, Belnap J, eds. Biological Soil Crusts: An Organizing Principle in Drylands. Cham, Switzerland: Springer, 257–285
|
| [5] |
Bates S T, Nash T H, Sweat K G, et al. 2010. Fungal communities of lichen-dominated biological soil crusts: Diversity, relative microbial biomass, and their relationship to disturbance and crust cover. Journal of Arid Environments, 74(10): 1192–1199
|
| [6] |
Belnap J. 1996. Soil surface disturbances in cold deserts: effects on nitrogenase activity in cyanobacterial-lichen soil crusts. Biology and Fertility of Soils, 23(4): 362–367. doi: 10.1007/BF00335908
|
| [7] |
Belnap J. 2002. Nitrogen fixation in biological soil crusts from southeast Utah, USA. Biology and Fertility of Soils, 35(2): 128–135. doi: 10.1007/s00374-002-0452-x
|
| [8] |
Belnap J. 2006. The potential roles of biological soil crusts in dryland hydrologic cycles. Hydrological Processes, 20(15): 3159–3178. doi: 10.1002/hyp.6325
|
| [9] |
Belnap J, Büdel B, Lange O L. 2001. Biological soil crusts: characteristics and distribution. In: Belnap J, Lange O L, eds. Biological Soil Crusts: Structure, Function, and Management. Berlin, Heidelberg: Springer, 3–30
|
| [10] |
Belnap J, Weber B, Büdel B. 2016. Biological soil crusts as an organizing principle in drylands. In: Weber B, Büdel B, Belnap J, eds. Biological Soil Crusts: An Organizing Principle in Drylands. Cham, Switzerland: Springer, 3–13
|
| [11] |
Beraldi-Campesi H, Hartnett H E, Anbar A, et al. 2009. Effect of biological soil crusts on soil elemental concentrations: implications for biogeochemistry and as traceable biosignatures of ancient life on land. Geobiology, 7(3): 348–359. doi: 10.1111/j.1472-4669.2009.00204.x
|
| [12] |
Bowker M A, Belnap J, Miller M E. 2006. Spatial modeling of biological soil crusts to support rangeland assessment and monitoring. Rangeland Ecology & Management, 59(5): 519–529
|
| [13] |
Brankatschk R, Fischer T, Veste M, et al. 2013. Succession of N cycling processes in biological soil crusts on a Central European inland dune. FEMS Microbiology Ecology, 83(1): 149–160. doi: 10.1111/j.1574-6941.2012.01459.x
|
| [14] |
Büdel B, Darienko T, Deutschewitz K, et al. 2009. Southern African biological soil crusts are ubiquitous and highly diverse in drylands, being restricted by rainfall frequency. Microbial Ecology, 57(2): 229–247. doi: 10.1007/s00248-008-9449-9
|
| [15] |
Castillo-Monroy A P, Bowker M A, Maestre F T, et al. 2011. Relationships between biological soil crusts, bacterial diversity and abundance, and ecosystem functioning: Insights from a semi-arid Mediterranean environment. Journal of Vegetation Science, 22(1): 165–174
|
| [16] |
Couradeau E, Giraldo-Silva A, De Martini F, et al. 2019. Spatial segregation of the biological soil crust microbiome around its foundational cyanobacterium, Microcoleus vaginatus, and the formation of a nitrogen-fixing cyanosphere. Microbiome, 7(1): 55. doi: 10.1186/s40168-019-0661-2
|
| [17] |
Darby B J, Neher D A. 2016. Microfauna within biological soil crusts. In: Weber B, Büdel B, Belnap J, eds. Biological Soil Crusts: An Organizing Principle in Drylands. Cham: Springer International Publishing, 139–157
|
| [18] |
Dixon R, Kahn D. 2004. Genetic regulation of biological nitrogen fixation. Nature Reviews Microbiology, 2(8): 621–631. doi: 10.1038/nrmicro954
|
| [19] |
Duan Zhenghu, Xiao Honglang. 2000. Effects of soil properties on ammonia volatilization. Soil Science and Plant Nutrition, 46(4): 845–852. doi: 10.1080/00380768.2000.10409150
|
| [20] |
Elbert W, Weber B, Burrows S, et al. 2012. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nature Geoscience, 5(7): 459–462
|
| [21] |
Evans R D, Ehleringer J R. 1993. A break in the nitrogen cycle in aridlands? Evidence from δ15N of soils. Oecologia, 94(3): 314–317. doi: 10.1007/BF00317104
|
| [22] |
Evans R D, Johansen J R. 1999. Microbiotic crusts and ecosystem processes. Critical Reviews in Plant Sciences, 18(2): 183–225. doi: 10.1080/07352689991309199
|
| [23] |
Evans R D, Lange O L. 2001. Biological soil crusts and ecosystem nitrogen and carbon dynamics. In: Belnap J, Lange O L, eds. Biological Soil Crusts: Structure, Function, and Management. Berlin, Heidelberg: Springer, 263–279
|
| [24] |
Fisher K A, Yarwood S A, James B R. 2017. Soil urease activity and bacterial ureC gene copy numbers: effect of pH. Geoderma, 285: 1–8. doi: 10.1016/j.geoderma.2016.09.012
|
| [25] |
Ghiloufi W, Seo J, Kim J, et al. 2019. Effects of biological soil crusts on enzyme activities and microbial community in soils of an arid ecosystem. Microbial Ecology, 77(1): 201–216. doi: 10.1007/s00248-018-1219-8
|
| [26] |
Guan Pingting, Zhang Xiaoke, Yu Jun, et al. 2018. Soil microbial food web channels associated with biological soil crusts in desertification restoration: the carbon flow from microbes to nematodes. Soil Biology and Biochemistry, 116: 82–90. doi: 10.1016/j.soilbio.2017.10.003
|
| [27] |
Hardy R W F, Burns R C, Holsten R D. 1973. Applications of the acetylene-ethylene assay for measurement of nitrogen fixation. Soil Biology and Biochemistry, 5(1): 47–81. doi: 10.1016/0038-0717(73)90093-X
|
| [28] |
He Zhili, Deng Ye, Van Nostrand J D, et al. 2010. GeoChip 3.0 as a high-throughput tool for analyzing microbial community composition, structure and functional activity. The ISME Journal, 4(9): 1167–1179. doi: 10.1038/ismej.2010.46
|
| [29] |
He Mei, Wang Wenxiong. 2011. Factors affecting the bioaccessibility of methylmercury in several marine fish species. Journal of Agricultural and Food Chemistry, 59(13): 7155–7162. doi: 10.1021/jf201424g
|
| [30] |
Hooper D U, Johnson L. 1999. Nitrogen limitation in dryland ecosystems: responses to geographical and temporal variation in precipitation. Biogeochemistry, 46(1): 247–293
|
| [31] |
Hu Yigang, Zhang Zhishan, Huang Lei, et al. 2019. Shifts in soil microbial community functional gene structure across a 61-year desert revegetation chronosequence. Geoderma, 347(2019): 126–134
|
| [32] |
Johnson S L, Budinoff C R, Belnap J, et al. 2005. Relevance of ammonium oxidation within biological soil crust communities. Environmental Microbiology, 7(1): 1–12. doi: 10.1111/j.1462-2920.2004.00649.x
|
| [33] |
Kruskal J B. 1964. Nonmetric multidimensional scaling: a numerical method. Psychometrika, 29(2): 115–129. doi: 10.1007/BF02289694
|
| [34] |
Kuypers M M, Marchant H K, Kartal B. 2018. The microbial nitrogen-cycling network. Nature Reviews Microbiology, 16(5): 263
|
| [35] |
Lan Shubin, Wu Li, Zhang Delu, et al. 2015. Analysis of environmental factors determining development and succession in biological soil crusts. Science of the Total Environment, 538: 492–499. doi: 10.1016/j.scitotenv.2015.08.066
|
| [36] |
Li Jie, Chen Qi, Zhang Si, et al. 2013. Highly heterogeneous bacterial communities associated with the SCS reef corals Porites lutea, Galaxea fascicularis and Acropora millepora. PLoS One, 8(8): e71301. doi: 10.1371/journal.pone.0071301
|
| [37] |
Ligi T, Truu M, Truu J, et al. 2014. Effects of soil chemical characteristics and water regime on denitrification genes (nirS, nirK, and nosZ) abundances in a created riverine wetland complex. Ecological Engineering, 72: 47–55. doi: 10.1016/j.ecoleng.2013.07.015
|
| [38] |
Liu Weiqiu, Song Yongsheng, Wang Bin, et al. 2012. Nitrogen fixation in biotic crusts and vascular plant communities on a copper mine tailings. European Journal of Soil Biology, 50: 15–20. doi: 10.1016/j.ejsobi.2011.11.009
|
| [39] |
Liu Yubin, Zhao Lina, Wang Zengru, et al. 2018. Changes in functional gene structure and metabolic potential of the microbial community in biological soil crusts along a revegetation chronosequence in the Tengger Desert. Soil Biology and Biochemistry, 126: 40–48. doi: 10.1016/j.soilbio.2018.08.012
|
| [40] |
Lorenzen C J. 1967. Determination of chlorophyll and phaeopigments: spectrophotometric equations. Limnology and Oceanography, 12(2): 343–346. doi: 10.4319/lo.1967.12.2.0343
|
| [41] |
Marker A F H, Jinks S. 1982. Spectrophotometric analysis of chlorophyll a and phaeopigments in acetone, ethanol and methanol. Ergebnisse der Limnologie, 16: 3–7
|
| [42] |
Miralles I, Domingo F, Cantón Y, et al. 2012. Hydrolase enzyme activities in a successional gradient of biological soil crusts in arid and semi-arid zones. Soil Biology and Biochemistry, 53: 124–132. doi: 10.1016/j.soilbio.2012.05.016
|
| [43] |
Mush E A. 1980. Comparison of different methods for chlorophyll and phaeopigment determination. Archiv für Hydrobiologie, Beihefte, 14: 14–36
|
| [44] |
Ochoa-Hueso R, Manrique E. 2011. Effects of nitrogen deposition and soil fertility on cover and physiology of Cladonia foliacea (Huds.) Willd., a lichen of biological soil crusts from Mediterranean Spain. Environmental Pollution, 159(2): 449–457. doi: 10.1016/j.envpol.2010.10.021
|
| [45] |
Orellana L H, Rodriguez-R L M, Higgins S, et al. 2014. Detecting nitrous oxide reductase (NosZ) genes in soil metagenomes: method development and implications for the nitrogen cycle. mBio, 5(3): e01193–14
|
| [46] |
Pointing S B, Belnap J. 2012. Microbial colonization and controls in dryland systems. Nature Reviews Microbiology, 10(8): 551–562. doi: 10.1038/nrmicro2831
|
| [47] |
Sivaperuman C, Jaisankar I, Velmurugan A, et al. 2008. Tropical islands: ecosystem and endemism. In: Sivaperuman C, Velmurugan A, Singh A K, et al., eds. Biodiversity and Climate Change Adaptation in Tropical Islands. Amsterdam: Elsevier, 31–52
|
| [48] |
Van Den Ancker J A M, Jungerius P D, Mur L R. 1985. The role of algae in the stabilization of coastal dune blowouts. Earth Surface Processes and Landforms, 10(2): 189–192. doi: 10.1002/esp.3290100210
|
| [49] |
Vitousek P M, Porder S, Houlton B Z, et al. 2010. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecological applications, 20(1): 5–15. doi: 10.1890/08-0127.1
|
| [50] |
Walvoord M A, Phillips F M, Stonestrom D A, et al. 2003. A reservoir of nitrate beneath desert soils. Science, 302(5647): 1021-1024
|
| [51] |
Wang Chao, Wang Xiaobo, Liu Dongwei, et al. 2014. Aridity threshold in controlling ecosystem nitrogen cycling in arid and semi-arid grasslands. Nature Communications, 5: 4799. doi: 10.1038/ncomms5799
|
| [52] |
Warshan D, Bay G, Nahar N, et al. 2016. Seasonal variation in nifH abundance and expression of cyanobacterial communities associated with boreal feather mosses. The ISME Journal, 10(9): 2198–2208. doi: 10.1038/ismej.2016.17
|
| [53] |
Weber B, Graf T, Bass M. 2012. Ecophysiological analysis of moss-dominated biological soil crusts and their separate components from the Succulent Karoo, South Africa. Planta, 236(1): 129–139. doi: 10.1007/s00425-012-1595-0
|
| [54] |
Wellburn A R. 1994. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Journal of Plant Physiology, 144(3): 307–313. doi: 10.1016/S0176-1617(11)81192-2
|
| [55] |
Yang Yunfeng, Gao Ying, Wang Shiping, et al. 2014. The microbial gene diversity along an elevation gradient of the Tibetan grassland. The ISME Journal, 8(2): 430–440. doi: 10.1038/ismej.2013.146
|
| [56] |
Yang Hangyu, Liu Changzhong, Liu Yanmei, et al. 2018. Impact of human trampling on biological soil crusts determined by soil microbial biomass, enzyme activities and nematode communities in a desert ecosystem. European Journal of Soil Biology, 87: 61–71. doi: 10.1016/j.ejsobi.2018.05.005
|
| [57] |
Yeager C M, Kuske C R, Carney T D, et al. 2012. Response of biological soil crust diazotrophs to season, altered summer precipitation, and year-round increased temperature in an arid grassland of the colorado plateau, USA. Frontiers in Microbiology, 3: 358
|
| [58] |
Yue Haowei, Wang Mengmeng, Wang Shiping, et al. 2015. The microbe-mediated mechanisms affecting topsoil carbon stock in Tibetan grasslands. The ISME Journal, 9(9): 2012–2020. doi: 10.1038/ismej.2015.19
|
| [59] |
Zehr J P, Paerl H. 1998. Nitrogen fixation in the marine environment: genetic potential and nitrogenase expression. In: Cooksey K E, ed. Molecular Approaches to the Study of the Ocean. Dordrecht: Springer, 285–301
|
| [60] |
Zhao Lina, Liu Yubing, Wang Zengru, et al. 2020. Bacteria and fungi differentially contribute to carbon and nitrogen cycles during biological soil crust succession in arid ecosystems. Plant and Soil, 447(1): 379–392
|
| [61] |
Zhou Jizhong, He Zhili, Yang Yunfeng, et al. 2015. High-throughput metagenomic technologies for complex microbial community analysis: open and closed formats. mBio, 6(1): e02288–14
|
| [62] |
Zumft W G. 1997. Cell biology and molecular basis of denitrification. Microbiology and Molecular Biology Review, 61(4): 533–616. doi: 10.1128/.61.4.533-616.1997
|
Figures(5) / Tables(7)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: