Yueyun Wang, Yadong Zhou, Chunsheng Wang. Ceuthonoe nezhai gen. et sp. n. (Polynoidae: Polynoinae) commensal with sponges from Weijia Guyot, western Pacific[J]. Acta Oceanologica Sinica, 2021, 40(12): 90-103. doi: 10.1007/s13131-021-1885-0
Citation:
Yueyun Wang, Yadong Zhou, Chunsheng Wang. Ceuthonoenezhai gen. et sp. n. (Polynoidae: Polynoinae) commensal with sponges from Weijia Guyot, western Pacific[J]. Acta Oceanologica Sinica, 2021, 40(12): 90-103. doi: 10.1007/s13131-021-1885-0
Yueyun Wang, Yadong Zhou, Chunsheng Wang. Ceuthonoe nezhai gen. et sp. n. (Polynoidae: Polynoinae) commensal with sponges from Weijia Guyot, western Pacific[J]. Acta Oceanologica Sinica, 2021, 40(12): 90-103. doi: 10.1007/s13131-021-1885-0
Citation:
Yueyun Wang, Yadong Zhou, Chunsheng Wang. Ceuthonoenezhai gen. et sp. n. (Polynoidae: Polynoinae) commensal with sponges from Weijia Guyot, western Pacific[J]. Acta Oceanologica Sinica, 2021, 40(12): 90-103. doi: 10.1007/s13131-021-1885-0
Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou 310012, China
2.
Key Laboratory of Marine Ecosystem Dynamics, Ministry of Natural Resources, Hangzhou 310012, China
3.
School of Oceanography, Shanghai Jiao Tong University, Shanghai 200030, China
4.
Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai), Zhuhai 519080, China
Funds:
The National Natural Science Foundation of China under contract No. 41806179; the China Ocean Mineral Resources Research and Development Association Program under contract No. DY135-E2-2-03.
Ceuthonoe, a new genus with type species C. nezhai sp. n. commensal with euplectellid glass sponge (Hexactinellida: Euplectellidae), is described and illustrated, based on the material collected from Weijia Guyot, western Pacific. The new genus is distinguished by a combination of characters: body with 32 segments; 14 pairs of elytra; prostomium with prominent cephalic peaks and facial tubercle; aciculae and chaetae on tentaculophores; biramous parapodium, each ramus with a long aciculum penetrating epidermis, without supra- or subacicular process; and noto- and neurochaetae with unidentate tips. Ceuthonoe gen. n. is mostly silimar to genera of Polynoinae in morphology. However the phylogenetic analysis based on cytochrome c oxidase subunit I, 16S, 18S, and 28S genes suggested that this new genus was separated from the main branch of Polynoinae. The study also discussed the genera related to the new genus.
Polynoidae is one of the most diverse polychaete families, in the number of genera and species (Salazar-Vallejo et al., 2015; Bonifácio and Menot, 2019). It is widely distributed from the intertidal zone to the hadal zone (Paterson et al., 2009). Polynoids are characterized by having dorsoventrally flattened bodies, two palps, one to three antennae, elytrae on the dorsum and biramous parapodia with simple chaetae. Among approximately 900 polynoid species, more than 200 are known to be involved in about 550 associations with other marine invertebrates (Britayev et al., 2014; Martin and Britayev, 2018).
To date, about 58 genera have been included in the subfamily Polynoinae (Read and Fauchald, 2019). Most primitive species in Polynoinae are obligate symbionts with specific morphological adaptations (Serpetti et al., 2017; Taboada et al., 2020). Eight species (Harmothoe hyalonemae Martín, Rosell & Uriz, 1992; H. spongicola Hanley & Burke, 1991; Hermadion fauveli Gravier, 1918; Hololepidella nigropunctata (Horst, 1915); Intoshella euplectellae (McIntosh, 1885); I. dictyaulus Sui, Li & Kou, 2018; Malmgreniella murrayensis Pettibone, 1993a; Neopolynoe acanellae (Verril, 1881)) from six genera in Polynoinae are commensal with sponges (Martin and Britayev, 1998, 2018). The host-specificity of polynoids is not very high, and some species may also be commensal with other hosts, such as corals.
In May 2016, a sponge specimen (Hexactinellida: Lyssacinosida: Euplectellidae) was collected by the Chinese manned deep-sea submersible Jiaolong at a depth of 2 505 m from Weijia Guyot, western Pacific. A pair of polynoids were separated from the inner cavity of the sponge. Following a detailed examination of the morphology and sequence comparisons of mitochondrial cytochrome c oxidase subunit I (COI) using the BLAST search in GenBank, a new species of Polynoinae was fully described and illustrated, and a new genus was proposed after morphological comparision and phylogenetic analysis.
2.
Materials and methods
Four gene fragments (COI, 16S, 18S, and 28S) were used to infer the phylogenetic relationship of the new specimens with other polynoid scale worms. The non-coding 16S, 18S, and 28S gene sequences were aligned separately with MAFFT v.7.450 (Katoh and Standley, 2013); COI gene sequences were aligned using the MUSCLE algorithm. The alignments were manually concatenated in Geneirous R.11, resulting in a concatenated alignment of 4 054 bp: COI (552 bp), 16S (520 bp), 18S (1 953 bp), and 28S (1 029 bp). Nucleotide substitution models were selected based on the BIC criterion using jModelTest v. 2.1.10: GTR + F + I + G4 for COI and 16S, TN+F+R3 for 18S and TIM3+F+R3 for 28S. Sigalionidae and Acoetidae were selected as the outgroups based on previous studies (Zhang et al., 2018; Taboada et al., 2020). Maximum likelihood (ML) analyses were conducted in IQtree 1.6.10, with the bootstrap support values for each node determined using an ultrafast bootstrap algorithm for 100 000 replicates. Bayesian inference (BI) analyses were implemented in MrBayes v3.2 (Ronquist et al., 2012), with the substitution models replaced by GTR+G+I. Two parallel chain runs were performed each for 2 000 000 generations, with topologies sampled at every 1 000 generations. Tracer v1.6 was used to estimate the burn-in value, visualize, and ensure convergence.
Two specimens were removed from the body cavity of the sponge (Hexactinellida: Lyssacinosida: Euplectellidae: Placopegma sp.) collected by submersible Jiaolong, from Weijia Guyot, the western Pacific in May 2016. The specimens were preserved in 95% ethanol, subsequently dissected, and examined under a stereomicroscope (Zeiss Discovery V20). Notochaetae and neurochaetae were dissected carefully from the body, observed and photographed under a microscope (Zeiss Axio Imager A2). For barcoding purposes, COI, 18S, 28S, and 16S genes were amplified and sequenced. DNA was extracted using a DNAeasy Tissue Kit (Qiagen) following the manufacturer’s protocol. Primer pairs and PCR programmes used to amplify polynoid genetic markers were the same as those described by Zhou et al. (2019). GenBank accession numbers are presented in Table 1. The type specimens are deposited in the Sample Repository of Second Institute of Oceanography (RSIO), Ministry of Natural Resources, Hangzhou, China.
Table
1.
List of taxa included in the phylogenetic analysis including the NCBI accession number
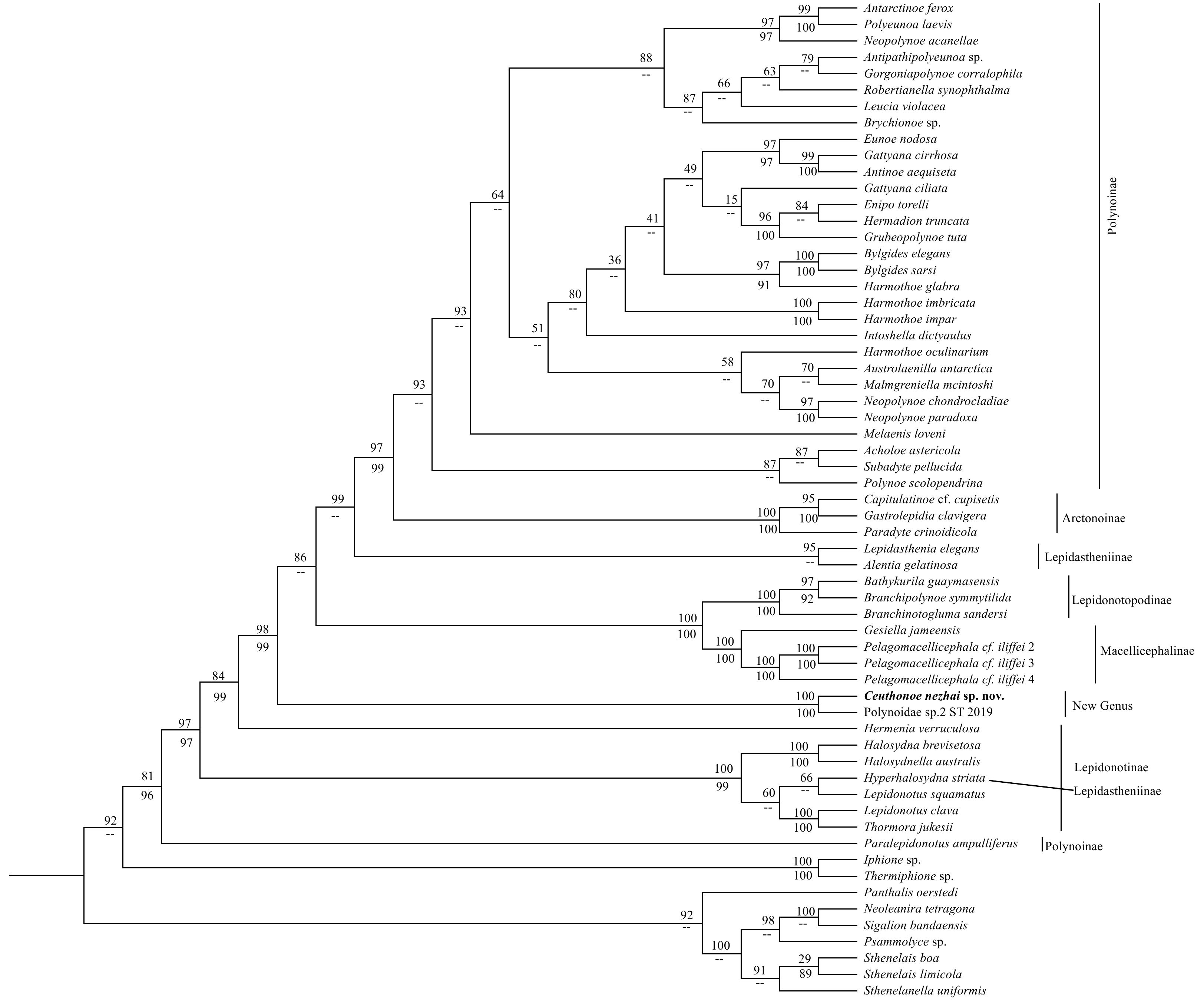
Pairwise GTR-corrected distances of COI between Ceuthonoe and non-Ceuthonoe genera, which averaged 27.50% (range 21.46%–33.74%), fell in the range of inter-generic genetic distance among non-Ceuthonoe genera (range 6.90%–38.31%, average 24.53%; Appendix Table A1). Both ML and BI analyses obtained similar topology, in which Ceuthonoenezhai sp. n. clustered with an undescribed polynoid commensal to a carnivorous sponge Chondrocladia verticillata Topsent, 1920, Polynoidae sp. 2 ST-2019 (Taboada et al., 2020), forming a discrete lineage outside a clade composed of species from Arctonoinae, Lepidastheniinae, Lepidonotopodinae, Macellicephalinae, and Polynoinae (Fig. 1).
Figure
1.
Phylogeny of polynoids based on the concatenated alignments of four genes using BI and ML analyses, imposed on the consensus ML tree. Node supports are shown as ML bootstrap values (BP, above) and Bayesian posterior probabilities (PP, below). Nodes not recovered in the BI analysis are marked with --.
Generic diagnosis. Body short, with about 32 segments. Elytra 14 pairs, on segments 2, 4, 5, and 7, alternate segments to 23, 26, and 29. Elytra without fringed papillae and with conical microtubercles.
Prostomium bilobed, with two distinct cephalic peaks, with paired palps, and three antennae. Lateral antennae with ceratophores inserted ventrally; a prominent conical facial tubercle under the median antenna. Eyes two pairs. Pharynx with two pairs of hooked jaws without denticulation on the inner border. Tentaculophore with an aciculum and few chaetae. Second segment with first pair of elytra and biramous parapodia.
Parapodia biramous; noto- and neuropodia well-developed with elongate acicular lobes; tips of noto- and neuroacicula penetrating the epidermis. Notochaetae thicker than neurochaetae. Nephridial papillae bulbous.
Etymology. The genus name is a combination of the Greek words “Ceuth” meaning “hidden” and “noe” from “Polynoe”, a sea nymph of Greek mythology. It refers to the type species commensal with sponges. Gender: feminine.
Remarks. This new genus is unique in having a low number of segments, prominent cephalic peaks, facial tubercle, aciculae, and chaetae on the tentacular segment, long aciculae spines penetrating the epidermis.
The new genus may be confused with Harmothoe, which has neurochaetae with bidentate tips whereas Ceuthonoe gen. n. has neurochaetae with unidentate tips. The new genus is also superficially similar to Antarctinoe Barnich et al. 2006, Bylgides Chamberlin 1919, Eunoe Malmgren 1866, and Gattyana McIntosh, 1897. They all have a low number of segments, cephalic peaks, and a facial tubercle. Antarctinoe and Bylgides are distinguishable from the new genus by having a digitiform supra-acicular process on neuropodia. The new genus differs from Eunoe and Gattyana in the form of chaetae. The distal tips of notochaetae of Eunoe are pointed and glabrous or blunt and split. Notochaetae of the new species have slightly curved, smooth tips. Neurochaetae of Eunoe are sturdy and have somewhat expanded and serrated subterminal parts, while that of the new genus are slender with tapering ends. Additionally, the neuropodia of Eunoe have a digitiform supra-acicular process (Wehe, 2006). Notochaetae of Gattyana are thinner than neurochaetae, while notochaetae of the new genus are thicker than neurochaetae.
Figure
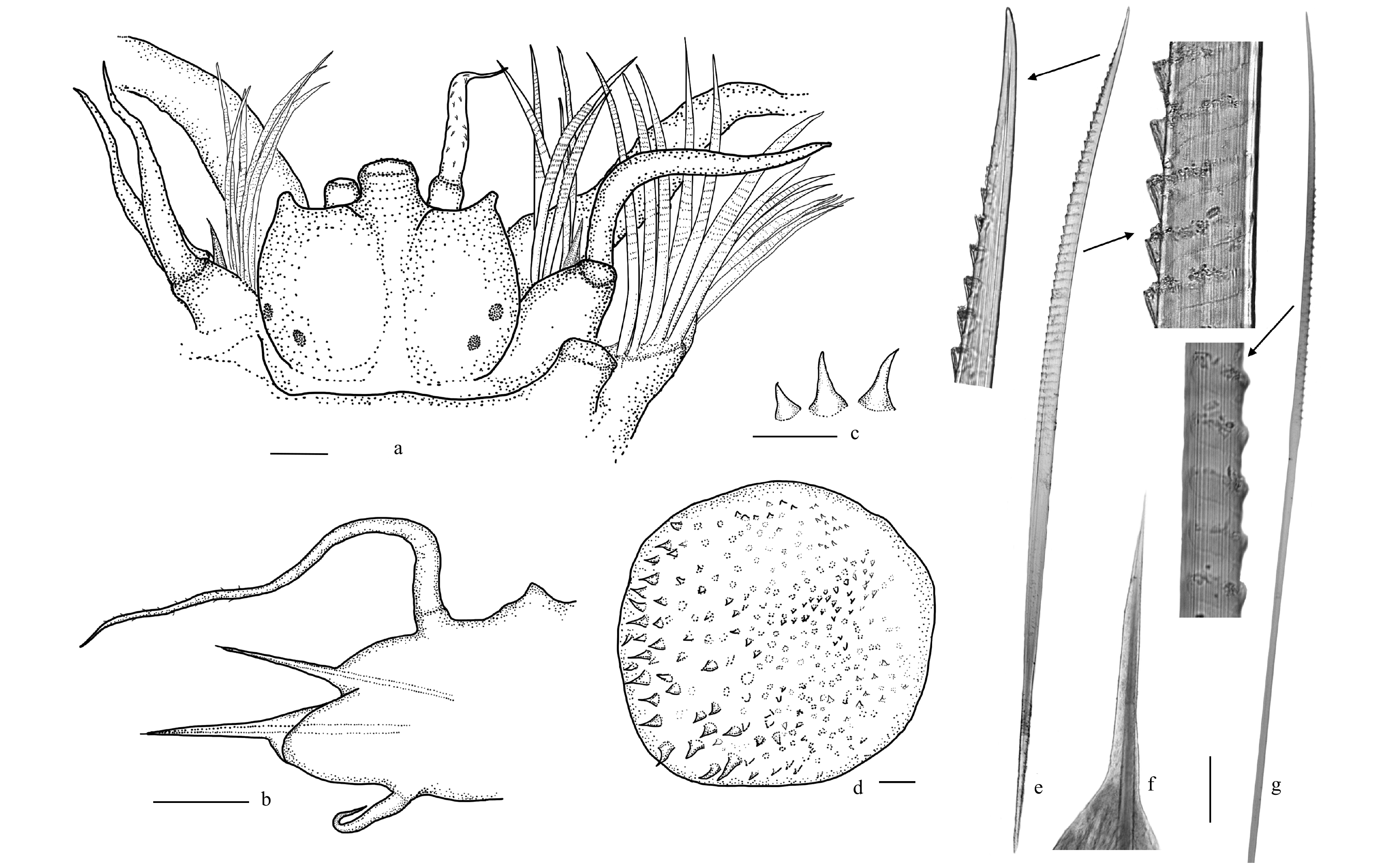
3.Ceuthonoe nezhai gen. et sp. n. Holotype. a. Prostomium, b. Parapodium of Segment 8 (chaetae omitted), end view, c. Tubercles on elytra, d. Elytra, e. Notochaeta and detailed view, f. Notoacicula, and g. Neurochaeta and detailed view. e–g are views under light microscope. Scale bars: 0.2 mm (a–d), 0.1 mm (e–g).
Type material. Holotype: B6317500002, complete, Weijia Guyot, western Pacific Ocean, JL-Dive106, Station S3-1, 13.018 45°N, 156.948 00°E, 2 505 m depth, May 1, 2016. Paratype: B6317500004, one complete specimen, same collecting data as holotype.
Description. Holotype about 19.8 mm from the prostomium to the end of the body (without anal cirri), 2.7 mm wide at the widest part excluding parapodia (Fig. 2a). Paratype about 15.2 mm in length and 1.8 mm wide at the widest part excluding parapodia (Fig. 2b). Parapodia long and subequal to the width of corresponding segments. The following description is based on holotype and paratype.
Body short, with 32 segments, tapering posteriorly; the ventral side flattened and the dorsal side slightly convex. A deep longitudinal groove on the ventral side. Elytra 14 pairs, on segments 2, 4, 5, and 7, alternate segments to 23, 26, and 29. Elytra large, covering the dorsum except the middle part. Elytra oval in shape, without fringed papillae, with many conical microtubercles on surfaces (Figs 2c, 3c and d). Microtubercles near anterior and inner regions increasing in number and becoming larger. Dorsal cirri and dorsal tubercles near each other, present on non-elytrigerous segments. Dorsal cirri composed of short cylindrical cirrophores and long styles, up to three times the length of the parapodia (Figs 2j and 3b).
Prostomium bilobed, wider than long, with distinct cephalic peaks and three antennae; ceratophore of median antenna in the anterior notch, lateral antennae with ceratophores inserted ventrally to median antenna (Figs 2d, e and 3a). Median antenna missing in both specimens. Styles of lateral antennae covered with short soft papillae (Fig. 2k). A pair of stout palps with smooth surface. A distinct conical facial tubercle under the ceratophore of the median antenna. Two pairs of eyes arranged in a reverse trapezium, posterolaterally on the widest part of the prostomium (Fig. 2f). Pharynx with two pairs of hooked jaws without denticulation on the inner border (Fig. 2l). Tentaculophores lateral to prostomium, each with a long projecting aciculum and many chaetae projecting anteriorly (Fig. 3a). Notochaetae similar to those in the following segments. Tentacular cirri shorter and thinner than palps.
Second segment with first elytra, biramous parapodia, and coiled ventral cirri longer than the following ones, without nuchal lobe. Parapodia biramous, with yellowish noto- and neuroaciculae and numerous chaetae (Figs 2m and 3b). Notopodium shorter than neuropodium, with a slender acicular lobe, and tip of aciculum penetrating epidermis (Fig. 3f). Notochaetae and neurochaetae the same from the anterior segments to the posterior end. Notochaetae stout, about 1.5 times as wide as neurochaetae. Notochaetae projecting dorsally, with numerous rows of spinules having slightly curved, smooth tips (Figs 2m and 3e). Neuropodium well developed, with obtuse postchaetal lobe and longer prechaetal acicular lobe, and tip of aciculum penetrating epidermis. The upper part of neurochaetae extending, wider than the shaft, with spinous regions. The tips of neurochaetae slightly curved and smooth (Fig. 3g). Ventral cirri short and less than half length of parapodia.
Nephridial papillae short bulbous, beginning on Segment 5. Anal cirri lost.
Pigmentation. Significant brown pigmentation on the ventral side of parapodia and body, especially on sides of the ventral groove. Tips of ventral cirri dark-brown. Pigmentation stable in alcohol-preserved specimens.
Type locality. Weijia Guyot, western Pacific.
Distribution. Only known from the type locality. Commensal with sponges.
Etymology. The specific name is dedicated to Nezha, which is a Chinese mythological figure.
Remarks. Polynoids are often found to be associated with other invertebrates, such as sponges, corals, echinoderms, mollusks, and other polychaetes. Eight Polynoinae species (Harmothoe hyalonemae from Western Mediterranean, 1 083–2 256 m; H. spongicola from Fairway Reefs, Coral Sea, 720 m; Hermadion fauveli from Cape Verde Islands, Atlantic Ocean; Hololepidella nigropunctata, widely distributed in Indo-west and central Pacific Ocean, 1–76 m; Intoshella euplectellae from Philippines, 184 m; I. dictyaulus from near the Mariana Trench, West Pacific, 890 m; Malmgreniella murrayensis from the South Arabian Coast, 38 m; Neopolynoe acanellae from east coast of the MA, USA) are reported commensal with sponges. Ceuthonoe nezhai sp. n. is similar to Harmothoe species in having distinct cephalic peaks, a facial tubercle, and lateral antennae inserted ventrally on the prostomium. However, the new species differs from the latter by possessing neurochaetae all with unidentate tips. Hololepidella, Intoshella and Malmgreniella species can also be easily distinguished from the new species by the number of segments, morphology of elytra, neurochaetae and prostomium. Hololepidella nigropunctata has 55 or more segments, with mostly smooth elytra, smooth dorsal cirri, and bidentate neurochaetae (Barnich et al., 2004; Wehe, 2006). Intoshella species has smooth elytra and no cephalic peaks. Malmgreniella murrayensis has 36 segments, no cephalic peaks, and bidentate neurochaetae. The new species is also closely related to species of Bylgides Chamberbin, 1919. They all have a small number of segments and elytrae, and aciculae on tentaculophores. They can be distinguished by the form of the chaetae and acicular lobes. Neurochaetae of Bylgides have long spinous regions and capillary tips, but the neurochaetae of Ceuthonoe nezhai sp. n. have blunt tips. Long conical neuroacicular lobes of Bylgides have a supraacicular digitiform process, but the acicular lobes of Ceuthonoe nezhai sp. n. have no supraacicular or subacicular process. The new species is also very similar to genus Neopolynoe species in anterior body. The species of Neopolynoe are distinguishable from the new species by having numerous segments and naked dorsum of the posterior body. Comparisons of diagnostic morphological characteristics of closely related genera with those of the new genus are presented in Table 2.
Table
2.
Morphological comparison of Genera related to Ceuthonoe gen. n. Unless otherwise indicated, character information is from original descriptions and illustrations
Polynoidae is one of the most diverse polychaetes family having the highest number of species involved in symbiotic relationships (Britayev et al., 2014). To date, phylogenetic relationships within Polynoidae have not been fully resolved. Phylogenetic analyses conducted on a limited number of polynoid sequences indicate that Polynoinae is not a monophyletic group (Bonifácio and Menot, 2019; Taboada et al., 2020). The morphological characteristics of the new genus suggest that it should be a member of Polynoinae based on the diagnosis given by Wehe (2006). However, the results obtained from molecular phylogenetic analyses reveal that Polynoinae is a polyphyletic group. The new genus and Paralepidonotus are separated from the major clade of Polynoinae, which is intermingled with species of Arctonoinae, Lepidastheniinae, Lepidonotopodinae, and Macellicephalinae in the phylogenetic tree, indicating that a wider range of taxa and genetic marker sampling is needed for further revision of this clade. Members belonging to Arctonoinae have subbiramous parapodia and no chaetae on tentaculophores, while those belonging to the Polynoinae usually have chaetae on tentaculophores and biramous parapodia (Barnich and Fiege, 2001). This new genus is more similar to a member of Polynoinae. Members of Lepidastheniinae are characterized by lateral antennae terminal, tentaculophores without chaetae, and reduced notopodia; notochaetae are usually missing (Salazar-Vallejo et al., 2015). The new genus is similar to Parahalosydna Horst, 1915 (Lepidastheniinae) in the number of segments and elytra. However, lateral antennae are in the terminal position in Parahalosydna and in the ventral position in the new genus. Subfamily Lepidonotopodinae was erected for genus Lepidonotopodium Pettibone (1983), characterized by having no lateral antennae. Bonifácio and Menot (2019) synonymized Lepidonotopodinae with Macellicephalinae based on the the phylogenetic analysis of morphological and molecular datasets. Both subfamilies were characterized by the synapomorphic absence of lateral antennae. Hatch et al. (2020) analyzed the phylogenetic relationships of Branchinotogluminae, Branchiplicatinae, Branchipolynoinae, Lepidonotopodinae, Bathykurila guaymasensis, and Levensteiniella (described as Macellicephalinae). The results suggested that these taxa formed a well-supported clade. Therefore, the oldest subfamily name Lepidonotopodinae Pettibone 1983 was reinstated, including Branchipolynoe, Branchinotogluma, Bathykurila, Branchiplicatus, Lepidonotopodium, Levensteiniella, Thermopolynoe, and Peinaleopolynoe. Our result showed that these two subfamilies were sister groups with robust support (Fig. 1). Lepidonotinae has a fixed number of segments (26) or more (up to 70 and more), with lateral antennae inserted terminally or subterminally on the prostomium. This new genus is closely related to the short-bodied genus Hermenia belonging to Lepidonotinae. Both of them have chaetae on the tentacular segment and a bulbous facial tubercle. Parapodia of Hermenia are subbiramous with reduced notopodia forming small lobes on the anterodorsal faces of stout neuropodia, while those of the new genus are biramous with normal-sized notopodia. Neurochaetae of Hermenia are stout, with falcate tips and 1 or 2 stout teeth. Neurochaetae of the new genus have unidentate tips. According to the taxonomic revision of Wehe (2006), the new genus is similar to the genera Eunoe and Harmothoe; they all have cephalic peaks, ventrally inserted lateral antennae, and chaetae on tentaculophores. Their differences are discussed in remarks on the new genus.
Symbiotic relationships between annelid polychaetes and other marine invertebrates are common; approximately 45% of all known commensal species belong to Polynoidae Kinberg, 1856 (Taboada et al., 2020). The species Polynoidae sp. 2 ST-2019 reported by Taboada et al. (2020) may also be included in this new genus owing to its close molecular phylogenetic relationship with Ceuthonoe nezhai gen. et sp. n. Both of them are similar in morphological characteristics. Polynoidae sp. 2 ST-2019 was collected in association with carnivorous sponge Chondrocladia verticillata from the Gulf of Mexico, while Ceuthonoe nezhai gen. et sp. n. was collected in association with sponge Placopegma sp. More findings are needed to clarify the aforementioned relationships.
Acknowledgements
We are very grateful to all the scientists and crew of the R/V Xiangyanghong 9 and the team of the submersible Jiaolong for their help: n collecting deep-sea materials. We thank Ruiyan Zhang for molecular data collection. We also thank Vlaams Instituut voor de Zee Library for their help in providing important references.
Barnich R, Fiege D. 2001. The Mediterranean species of Malmgreniella Hartman, 1967 (Polychaeta: Polynoidae: Polynoinae), including the description of a new species. Journal of Natural History, 35(8): 1119–1142. doi: 10.1080/00222930152434463
[2]
Barnich R, Fiege D, Micaletto G, et al. 2006. Redescription of Harmothoe spinosa Kinberg, 1856 (Polychaeta: Polynoidae) and related species from Subantarctic and Antarctic waters, with the erection of a new genus. Journal of Natural History, 40(1–2): 33–75. doi: 10.1080/00222930500445044
[3]
Barnich R, Fiege D, Sun Ruiping. 2004. Polychaeta (Annelida) of Hainan Island, South China Sea: Part III. Aphroditoidea. Species Diversity, 9(4): 285–329. doi: 10.12782/specdiv.9.285
[4]
Bock G, Fiege D, Barnich R. 2010. Revision of Hermadion Kinberg, 1856, with a redescription of Hermadion magalhaensi Kinberg, 1856, Adyte hyalina (G. O. Sars, 1873) n. comb. and Neopolynoe acanellae (Verrill, 1881) n. comb. (Polychaeta: Polynoidae). Zootaxa, 2554: 45–61. doi: 10.11646/zootaxa.2554.1.4
[5]
Bonifácio P, Menot L. 2019. New genera and species from the Equatorial Pacific provide phylogenetic insights into deep-sea Polynoidae (Annelida). Zoological Journal of the Linnean Society, 185(3): 555–635. doi: 10.1093/zoolinnean/zly063
[6]
Britayev T A, Gil J, Altuna Á, et al. 2014. New symbiotic associations involving polynoids (Polychaeta, Polynoidae) from Atlantic waters, with redescriptions of Parahololepidella greeffi (Augener, 1918) and Gorgoniapolynoe caeciliae (Fauvel, 1913). Memoirs of Museum Victoria, 71: 27–43. doi: 10.24199/j.mmv.2014.71.04
[7]
Fiege D, Barnich R. 2009. Polynoidae (Annelida: Polychaeta) associated with cold-water coral reefs of the northeast Atlantic and the Mediterranean Sea. Zoosymposia, 2: 149–164. doi: 10.11646/zoosymposia.2.1.13
[8]
Jirkov I A. 2001. Polikhety severnogo Ledovitogo Okeana [Polychaeta of the Arctic Ocean] (in Russian). Moscow, Russia: Yanus-K Press, 126–127
[9]
Hanley J R. 1991. Revision of the genus Parlalepidonotus Horst, 1915 (Polychaeta: Polynoidae). Invertebrate Taxonomy, 4(5): 1053–1075
[10]
Hanley J R. 1993. Scaleworms (Polychaeta: Polynoidae) of Rottnest Island, Western Australia. In: Wells F E, Walker D I, Kirkman H, et al., eds. Proceedings of the Fifth International Marine Biological Workshop: The Marine Flora and Fauna of Rottnest Island, Western Australia. Perth: Western Australian Museum, 305–320
[11]
Hanley J R, Burke M. 1991. Polychaeta Polynoidae: Scaleworms of the Chesterfield Islands and Fairway Reefs, Coral Sea. In: Crosnier A, ed. Résultats des Campagnes MUSORSTOM, Volume 8. Mémoires du Muséum national d'histoire naturelle, 151(1): 9–82
[12]
Hatch A S, Liew H, Hourdez S, et al. 2020. Hungry scale worms: Phylogenetics of Peinaleopolynoe (Polynoidae, Annelida), with four new species. ZooKeys, 932: 27–74. doi: 10.3897/zookeys.932.48532
[13]
Katoh K, Standley D M. 2013. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Molecular Biology and Evolution, 30(4): 772–780. doi: 10.1093/molbev/mst010
[14]
Levenstein R Y. 1982. New genera of the subfamily macellicephalinae (Polychaeta, Polynoidae) from the Tasman hollow. Zoologicheskii Zhurnal, 61(9): 1291–1296
[15]
Martin D, Britayev T A. 1998. Symbiotic polychaetes: review of known species. Oceanography and Marine Biology: An Annual Review, 36: 217–340
[16]
Martin D, Britayev T A. 2018. Symbiotic polychaetes revisited: An update of the known species and relationships (1998–2017). Oceanography and Marine Biology, 56: 371–448
[17]
Martín D, Rosell D, Uriz M J. 1992. Harmothöe hyalonemae sp. nov. (Polychaeta, Polynoidae), an exclusive inhabitant of different Atlanto-Mediterranean species of Hyalonema (Porifera, Hexactinellida). Ophelia, 35(3): 169–185. doi: 10.1080/00785326.1992.10429925
[18]
McIntosh W C. 1885. Report on the Annelida Polychaeta collected by H. M. S. Challenger during the years 1873–76. Report on the Scientific Results of the Voyage of H. M. S. Challenger during the years 1873–76. Zoology, 12: 1–554
[19]
Paterson G L J, Glover A G, Barrio Froján C R S, et al. 2009. A census of abyssal polychaetes. Deep Sea Research Part II: Topical Studies in Oceanography, 56(19–20): 1739–1746. doi: 10.1016/j.dsr2.2009.05.018
[20]
Pettibone M H. 1975. Review of the genus Hermenia, with a description of a new species (Polychaeta: Polynoidae: Lepidonotinae). Proceedings of the Biological Society of Washington, 88(22): 233–248
[21]
Pettibone M H. 1983. A new scale worm (Polychaeta: Polynoidae) from the hydrothermal rift-area off western Mexico at 21°N. Proceedings of the Biological Society of Washington, 96(3): 392–399
[22]
Pettibone M H. 1993a. Scaled polychaetes (Polynoidae) associated with ophiuroids and other invertebrates and review of species referred to Malmgrenia McIntosh and replaced by Malmgreniella Hartman, with descriptions of new taxa. Smithsonian Contributions to Zoology, 538: 1–92
[23]
Pettibone M H. 1993b. Revision of some species referred to Antinoe, Antinoella, Antinoana, Bylgides, and Harmothoe (Polychaeta: Polynoidae: Harmothoinae). Smithsonian Contributions to Zoology, 545: 1–41
[24]
Read G, Fauchald K. 2019. World Polychaeta database. Polynoidae Kinberg, 1856. World Register of Marine Species. http://www.marinespecies.org/aphia.php?p=taxdetails&id=939 [2019-12-30/2021-04-01]
[25]
Ronquist F, Teslenko M, Van der Mark P, et al. 2012. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology, 61(3): 539–542. doi: 10.1093/sysbio/sys029
[26]
Salazar-Vallejo S I, González N E, Salazar-Silva P. 2015. Lepidasthenia loboi sp. n. from Puerto Madryn, Argentina (Polychaeta, Polynoidae). ZooKeys, 546: 21–37. doi: 10.3897/zookeys.546.6175
[27]
Serpetti N, Taylor M L, Brennan D, et al. 2017. Ecological adaptations and commensal evolution of the Polynoidae (Polychaeta) in the Southwest Indian Ocean Ridge: A phylogenetic approach. Deep Sea Research Part II: Topical Studies in Oceanography, 137: 273–281. doi: 10.1016/j.dsr2.2016.06.004
[28]
Sui Jixing, Li Xinzheng, Kou Qi. 2019. A new species of the genus Intoshella Darboux, 1899 (Polychaeta: Polynoidae) commensal with a deep-sea sponge from a seamount near the Mariana Trench. Marine Biodiversity, 49(3): 1479–1488. doi: 10.1007/s12526-018-0922-5
[29]
Taboada S, Silva A S, Neal L, et al. 2020. Insights into the symbiotic relationship between scale worms and carnivorous sponges (Cladorhizidae, Chondrocladia). Deep Sea Research Part I: Oceanographic Research Papers, 156: 103191. doi: 10.1016/j.dsr.2019.103191
[30]
Uschakov P V. 1982. Polychaetes of the Suborder Aphroditiformia of the Arctic Ocean and the Northwestern Part of the Pacific, Families Aphroditidae and Polynoidae. Fauna of the USSR, Polychaeta, 2: 1–272
[31]
Wehe T. 2006. Revision of the scale worms (Polychaeta: Aphroditoidea) occurring in the seas surrounding the Arabian Peninsula: Part I. Polynoidae. Fauna of Arabia, 22: 23–197
[32]
Zhang Yanjie, Sun Jin, Rouse G W, et al. 2018. Phylogeny, evolution and mitochondrial gene order rearrangement in scale worms (Aphroditiformia, Annelida). Molecular Phylogenetics and Evolution, 125: 220–231. doi: 10.1016/j.ympev.2018.04.002
[33]
Zhou Yadong, Chen Chong, Sun Yanan, et al. 2019. Amphisamytha (Annelida: Ampharetidae) from Indian Ocean hydrothermal vents: Biogeographic implications. Deep Sea Research Part I: Oceanographic Research Papers, 154: 103148. doi: 10.1016/j.dsr.2019.103148
Yuru Han, Yadong Zhou, Chong Chen, et al. Diversity and biogeography of scale worms in the subfamily Lepidonotopodinae (Annelida: Polynoidae) from Indian Ocean hydrothermal vents with descriptions of four new species. Zoological Journal of the Linnean Society, 2023. doi:10.1093/zoolinnean/zlad140
Yueyun Wang, Yadong Zhou, Chunsheng Wang. Ceuthonoe nezhai gen. et sp. n. (Polynoidae: Polynoinae) commensal with sponges from Weijia Guyot, western Pacific[J]. Acta Oceanologica Sinica, 2021, 40(12): 90-103. doi: 10.1007/s13131-021-1885-0
Yueyun Wang, Yadong Zhou, Chunsheng Wang. Ceuthonoenezhai gen. et sp. n. (Polynoidae: Polynoinae) commensal with sponges from Weijia Guyot, western Pacific[J]. Acta Oceanologica Sinica, 2021, 40(12): 90-103. doi: 10.1007/s13131-021-1885-0
Table
2.
Morphological comparison of Genera related to Ceuthonoe gen. n. Unless otherwise indicated, character information is from original descriptions and illustrations
Figure 1. Phylogeny of polynoids based on the concatenated alignments of four genes using BI and ML analyses, imposed on the consensus ML tree. Node supports are shown as ML bootstrap values (BP, above) and Bayesian posterior probabilities (PP, below). Nodes not recovered in the BI analysis are marked with --.
Figure 2. Ceuthonoe nezhai gen. et sp. n.: Holotype, dorsal view (a), Paratype, ventral view (b), Elytra from Segment 2 (c), Appendages of prostomium from holotype and paratype (d and e), Dorsal view of prostomium (first pair of elytra removed) (f) Median segments, ventral view (g), anal segment, dorsal and ventral view (h and i), Dorsal cirrus (j), Lateral antenna (k), Dorsal view of jaws from paratype (l), and Parapodium of Segment 8, end view (m). dc: dorsal cirrus, dtc: dorsal tentacular cirrus, ep: eltrophore, ft: facial tubercle, laa: lateral antenna, lac: lateral antenna ceratophore, mac: median antenna ceratophore, np: nephridial papilla, pal: palp, vc: ventral cirrus, vtc: ventral tentacular cirrus, 2vc: ventral cirrus on Segment 2, and 3vc: ventral cirrus on Segment 3. Scale bars: 2.0 mm (a, b, k), 0.2 mm (c–e), 0.5 mm (f–j), and 1.0 mm (m).
Figure 3. Ceuthonoe nezhai gen. et sp. n. Holotype. a. Prostomium, b. Parapodium of Segment 8 (chaetae omitted), end view, c. Tubercles on elytra, d. Elytra, e. Notochaeta and detailed view, f. Notoacicula, and g. Neurochaeta and detailed view. e–g are views under light microscope. Scale bars: 0.2 mm (a–d), 0.1 mm (e–g).



 DownLoad:
DownLoad:
 DownLoad:
DownLoad: