College of Life and Geographic Sciences, Key Laboratory of Biological Resources and Ecology of Pamirs Plateau in Xinjiang Uygur Autonomous Region, Kashi University, Kashi 844000, China
2.
Institute of Evolution & Marine Biodiversity, Ocean University of China, Qingdao 266003, China
Funds:
Tthe National Natural Science Foundation of China under contact No.32270434
All vertebrates have a definitive thyroid gland, or thyroid for short. As a critical organ for growth, development and metabolism, its origin and evolution have long received attention. On the basis of anatomical position, endodermal origination and histological features, the endostyle of amphioxus has been proposed as a homologue to the thyroid of vertebrates. This homology is further supported by the findings that the amphioxus endostyle abounds in thyroid hormones, possesses several thyroid-specific proteins such as thyroperoxidase, NADPH oxidase and thyroglobulin, and expresses the thyroid-related transcription factors involved in the regulation of development of the vertebrate thyroid, including Nkx2.1, FoxE4 and Pax2/5/8. Importantly, our study on functionality, together with others, indicates significant similarities between the amphioxus endostyle and the vertebrate thyroid gland. Moreover, we show that the production of thyroid hormones by the amphioxus endostyle is mediated in a fashion similar to that of the vertebrate thyroid. These provide solid evidences that the amphioxus endostyle is the homologue of the vertebrate thyroid. From a phylogenetic viewpoint, we propose that the hypobranchial ridge, or endostyle-like structure, of hemichordates is the most primitive forerunner of the thyroid, from which the vertebrate thyroid is formed through the transformation of non-follicular endostyle of amphioxus to follicular endocrine organ of vertebrates. We also raise a couple of questions that demand further study.
Figure 1. Views of phylogenic relationships of deuterostomes and evolution of chordates. Ascidian (urochordate) was previously regarded as the basal chordate, which evolved first, and then amphioxus (cephalochordate) and vertebrates. The positions of ascidian and amphioxus are now reversed by the results of the studies of molecular biology and developmental genetics, placing ascidian as sister group of the vertebrates and amphioxus basally in the chordates.
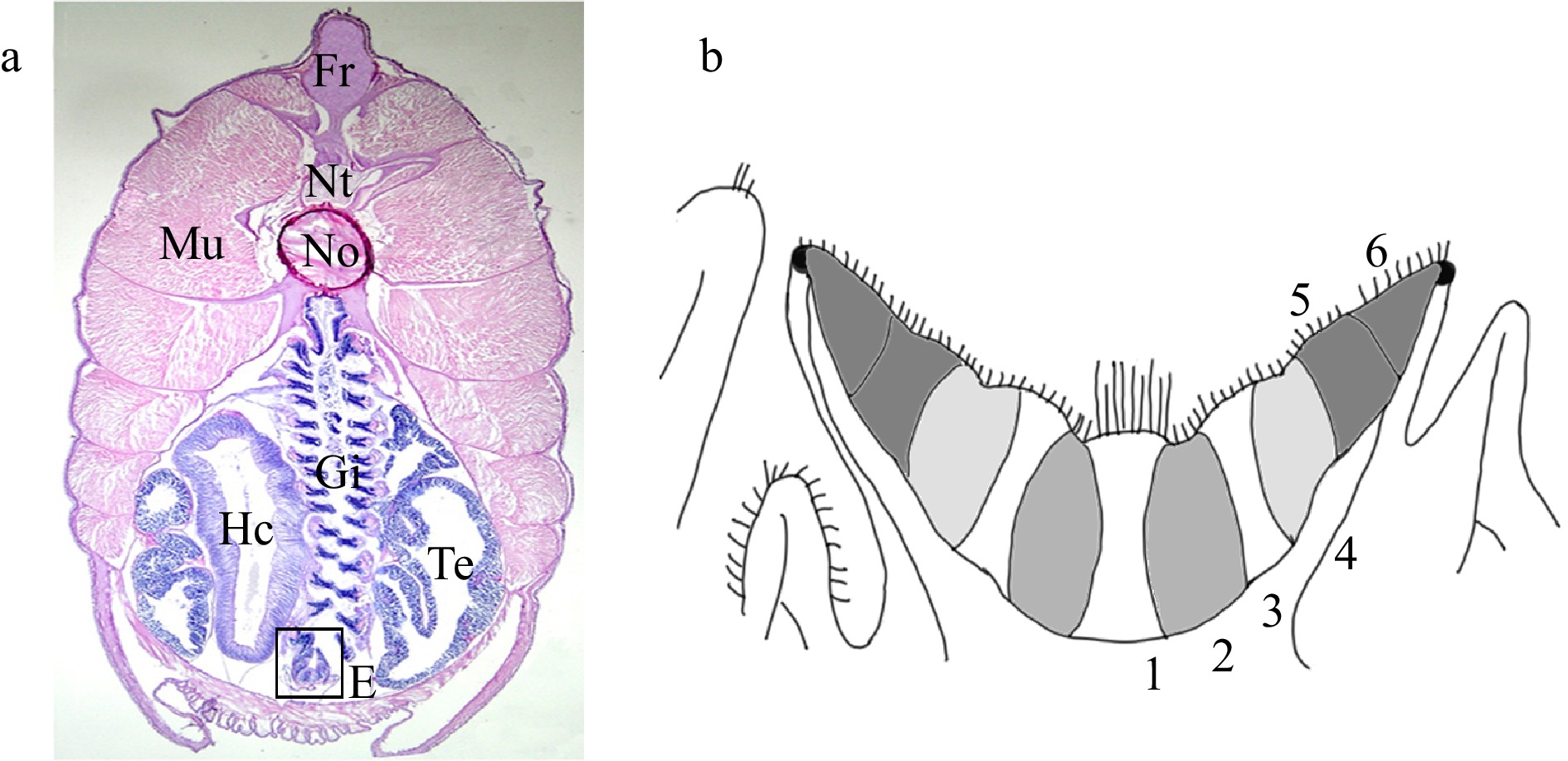
Figure 2. Endostyle of amphioxus. a. A cross section stained with H and E, showing the relative position of endostyle (box). Fr, fin ray; Hc, hepatic caecum; Mu, muscle; No, notochord; Nt, neural tube; Gi, gill; Te, testis; E, endostyle. b. A diagram of amphioxus endostyle. The midline of the endostyle is indicated by dotted lines. The endostyle consists of six zones. Zone 1 occupies the base of endostylar groove. Zones 2 to 6 occupy the lateral walls of endostyle. On the apical surface of cells of zone 1, long cilia exist, especially in the anterior endostyle. Cells of zones 2 to 6 have relatively short cilia. Cells of zones 1, 3, 5, and 6 are densely packed with flattened nuclei, whereas in cells of zones 2 and 4 the nuclei are restricted to the base.
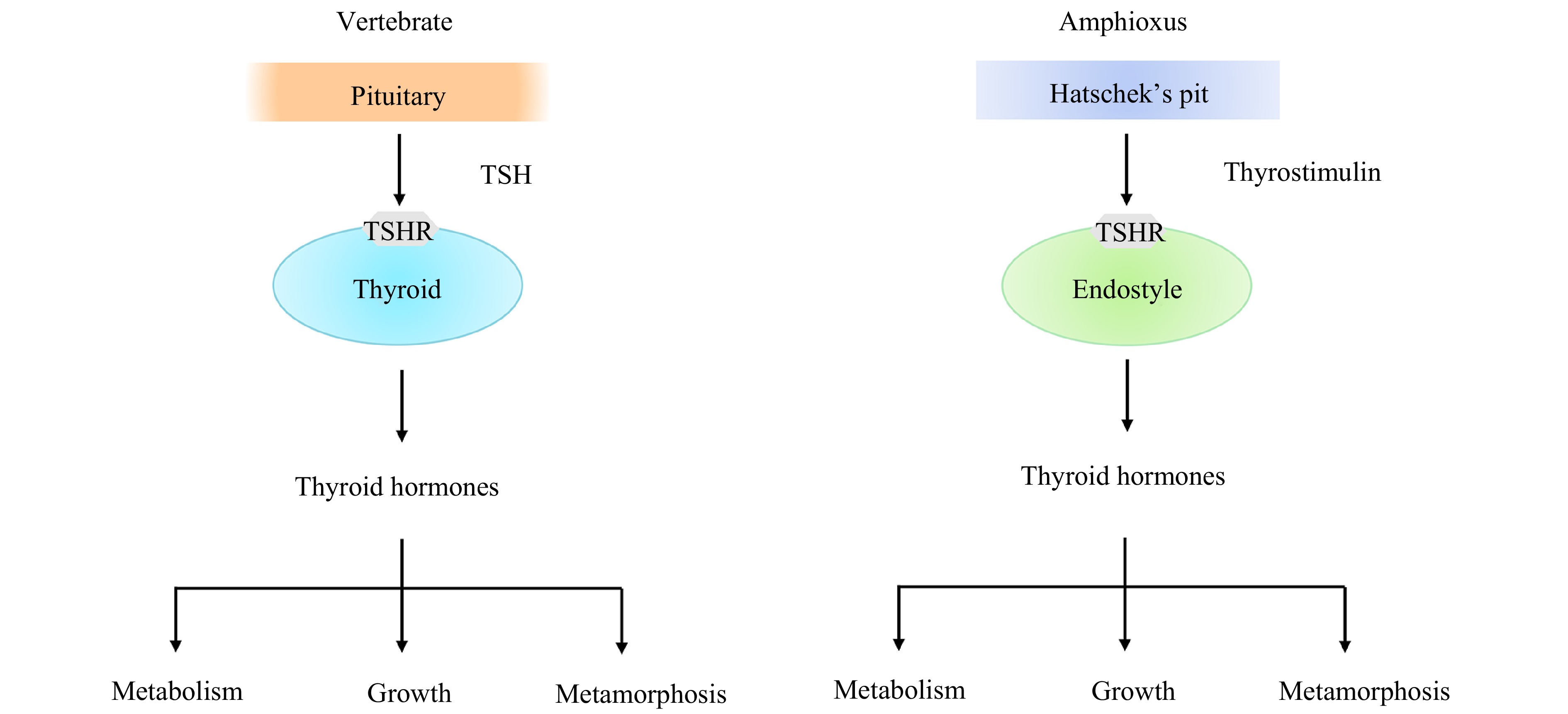
Figure 3. A diagram showing regulation of TH production and functions of THs in amphioxus and vertebrates. In vertebrates, the production of THs in the thyroid is mediated via the binding of the pituitary hormone TSH to the receptor TSHR, while in amphioxus, the production of THs in the endostyle is mediated via the binding of the thyrostimulin of Hatschek’s pit to the receptor TSHR.
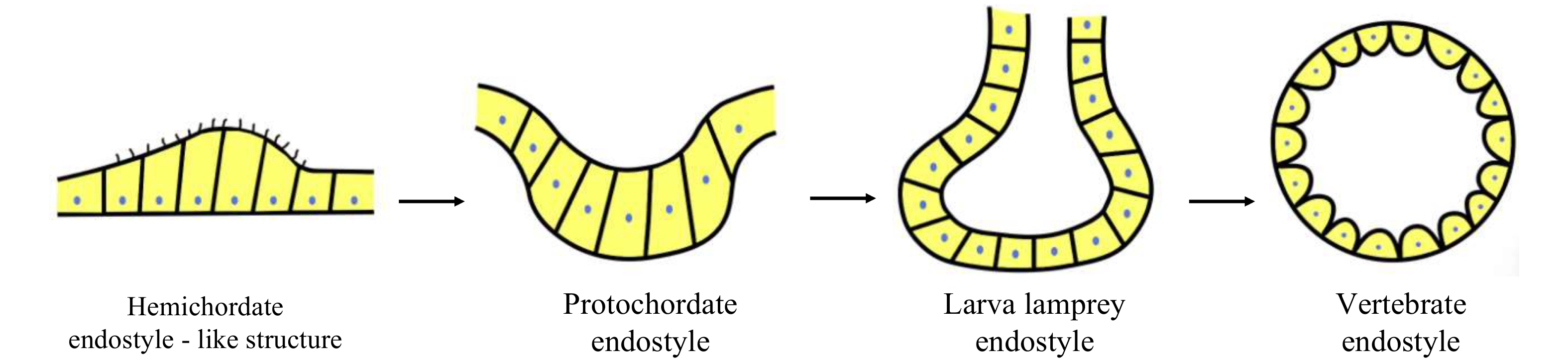
Figure 4. A diagram showing the hypothesis about origin and evolution of the vertebrate thyroid gland. The hemichordate endostyle is the most primitive forerunner of the vertebrate thyroid. The evolution of the vertebrate thyroid involves the transformation of non-follicular endostyle typical of the endostyle of the protochordates (amphioxus and ascidian) and larval lampreys to follicular endocrine organ characteristic of the thyroid of adult lampreys and jawed vertebrates.








 DownLoad:
DownLoad: