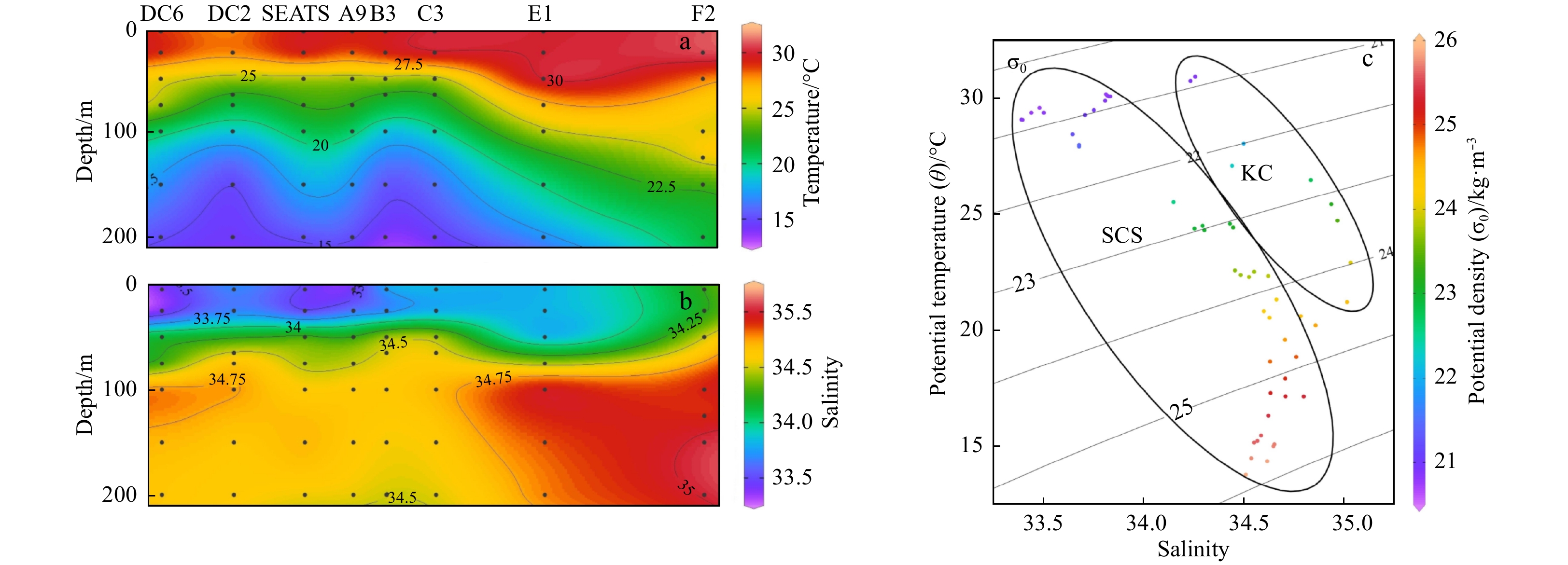
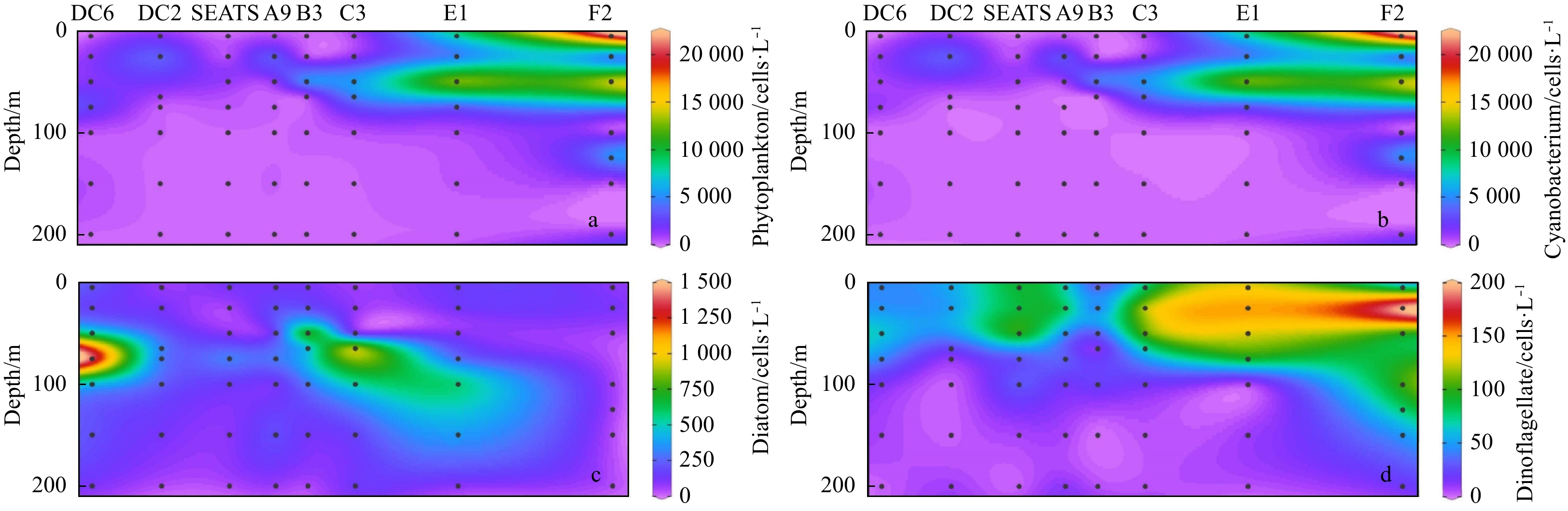
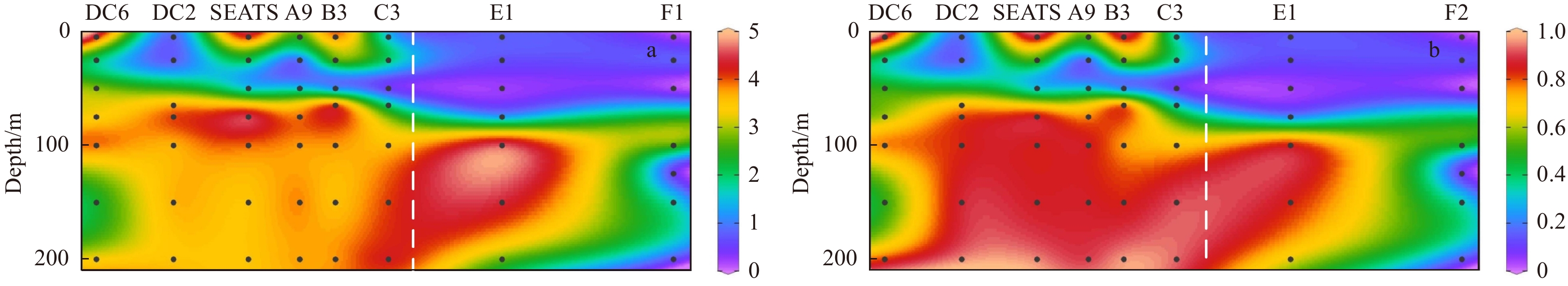
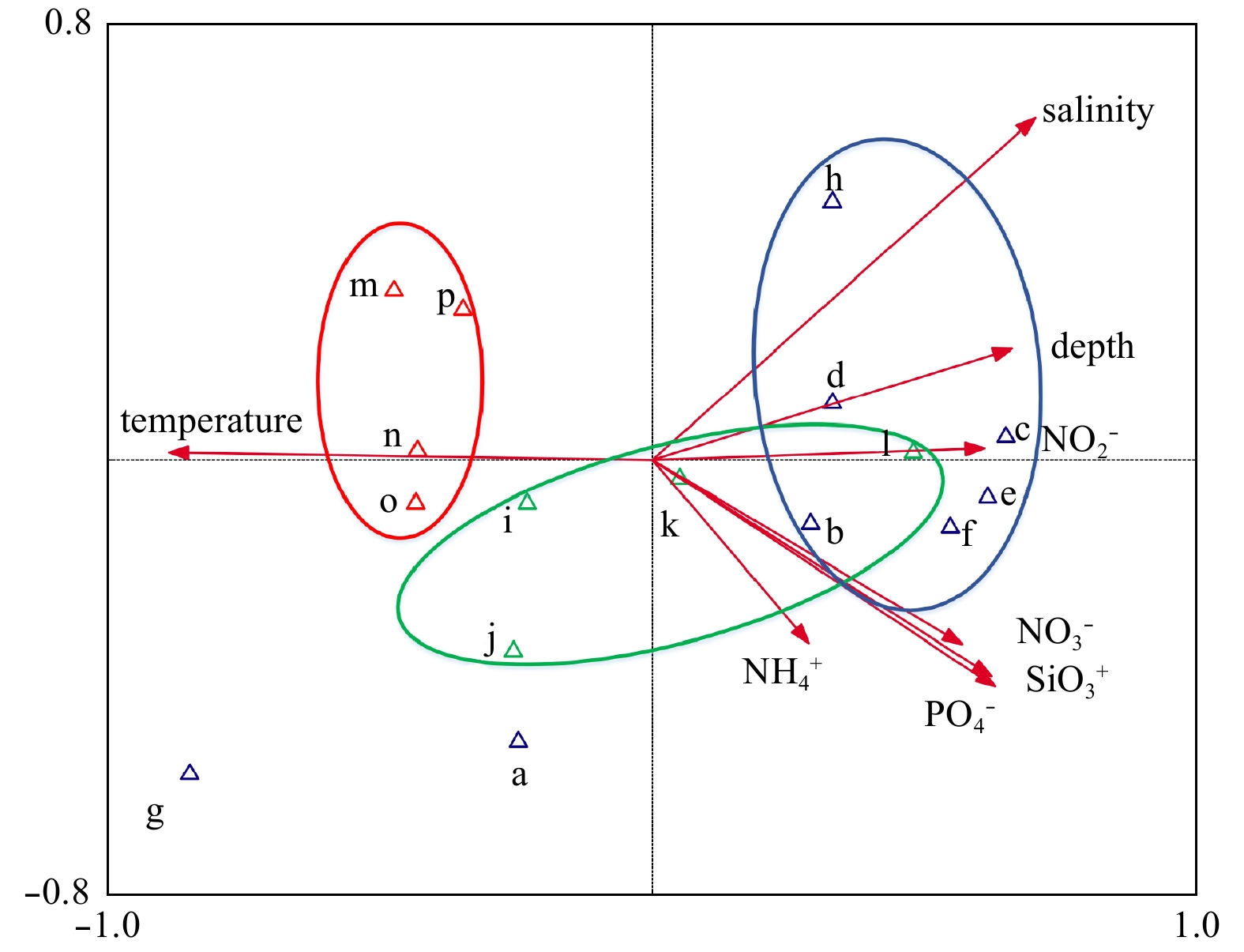
| Citation: | Xingzhou Wang, Yuqiu Wei, Chao Wu, Congcong Guo, Jun Sun. The profound influence of Kuroshio intrusion on microphytoplankton community in the northeastern South China Sea[J]. Acta Oceanologica Sinica, 2020, 39(8): 79-87. doi: 10.1007/s13131-020-1608-y

|

| [1] |
Caruso M J, Gawarkiewicz G G, Beardsley R C. 2006. Interannual variability of the Kuroshio intrusion in the South China Sea. Journal of Oceanography, 62(4): 559–575. doi: 10.1007/s10872-006-0076-0
|
| [2] |
Centurioni L R, Niiler P P, Lee D K. 2004. Observations of inflow of Philippine Sea surface water into the South China Sea through the Luzon Strait. Journal of Physical Oceanography, 34: 113–121. doi: 10.1175/1520-0485(2004)034<0113:OOIOPS>2.0.CO;2
|
| [3] |
Chen Y F L. 2005. Spatial and seasonal variations of nitrate-based new production and primary production in the South China Sea. Deep-Sea Research Part I: Oceanographic Research Papers, 52(2): 319–340. doi: 10.1016/j.dsr.2004.11.001
|
| [4] |
Chen Gengxin, Hu Po, Hou Yijun, et al. 2011. Intrusion of the Kuroshio into the South China Sea, in September 2008. Journal of Oceanography, 67(4): 439–448. doi: 10.1007/s10872-011-0047-y
|
| [5] |
Dai Minghan, Wang Lei, Guo Xianghui, et al. 2008. Nitrification and inorganic nitrogen distribution in a large perturbed river/estuarine system: the Pearl River Estuary, China. Biogeosciences, 5: 1227–1244. doi: 10.5194/bg-5-1227-2008
|
| [6] |
Ding Changling. 2009. The abundance, the spatial and temporal distributions of cyanobacteria in the Yellow Sea, the East China Sea and the South China Sea (in Chinese) [dissertation]. Qingdao: Ocean University of China
|
| [7] |
Fujioka S. 1990. Illustrations of the plankton of the Kuroshio-waters. Plankton in Amami-oshima Island Coastal Waters. Nagasaki, Japan: Nagasaki-ken Culture Publishing Union
|
| [8] |
Guo Shujing, Feng Yuanyuan, Wang Lei, et al. 2014. Seasonal variation in the phytoplankton community of a continental-shelf sea: The East China Sea. Marine Ecology Progress, 516: 103–126. doi: 10.3354/meps10952
|
| [9] |
Isamu Y. 1962. Illustrations of the Marine Plankton of Japan. Tokyo: Hoikusha Publishing
|
| [10] |
Jaccard P. 1908. Nouvelles recherches sur la distribution florale. Bulletin de la Societe Vaudoise des Sciences Naturelles, 44: 223–270
|
| [11] |
Jin Dexiang. 1965. Chinese Marine Planktonic Diatoms (in Chinese). Shanghai: Shanghai Scientific and Technical Publishers
|
| [12] |
Ke Zhixin, Huang Liangmin, Tan Yehui, et al. 2011. Species composition and abundance of phytoplankton in the northern South China Sea in summer 2007. Journal of Tropical Oceanography (in Chinese), 30(1): 131–143
|
| [13] |
Le Fengfeng, Sun Jun, Ning Xiuren, et al. 2006. Phytoplankton in the northern South China Sea in summer 2004. Oceanologia et Limnologia Sinica (in Chinese), 37(3): 238–248
|
| [14] |
Li Xin, Sun Jun, Tian Wei, et al. 2012. Phytoplankton community in the northern area of South China Sea in summer of 2009. Marine Sciences (in Chinese), 36(10): 33–39
|
| [15] |
Li Ruixiang, Zhu Mingyuan, Wang Zongling, et al. 2003. Mesocosm experiment on competition between two HAB species in East China Sea. Chinese Journal of Applied Ecology (in Chinese), 14(7): 1049–1054
|
| [16] |
Liu Dongyan, Sun Jun, Liu Zhe, et al. 2004. The effects of spring-neap tide on the phytoplankton community development in the Jiaozhou Bay, China. Acta Oceanologica Sinica, 23(4): 687–697
|
| [17] |
Ma Wei, Sun Jun. 2014. Characteristics of phytoplankton community in the northern South China Sea in summer and winter. Acta Ecologica Sinica (in Chinese), 34(3): 621–632
|
| [18] |
Pai Sucheng, Tsau Y J, Yang T I. 2001. pH and buffering capacity problems involved in the determination of ammonia in saline water using the indophenol blue spectrophotometric method. Analytica Chimica Acta, 434(2): 209–216. doi: 10.1016/S0003-2670(01)00851-0
|
| [19] |
Pielou E C. 1969. An introduction to mathematical ecology. New York: Wiley Interscience
|
| [20] |
Shannon C, Weaver W. 1949. The Mathematical Theory of Communication. Urbana IL: University of Illinois Press
|
| [21] |
Sun Ruyong. 1987. The Principle of Animal Ecology (in Chinese). Beijing: Beijing Normal University Press
|
| [22] |
Sun Jun. 2011. Marine phytoplankton and biological carbon sink. Acta Ecologica Sinica (in Chinese), 31(18): 5372–5378
|
| [23] |
Sun Jian, Hou Lipei, Xie Julun. 2006. Preliminary analysis of seasonal variation of Kuroshio in Luzon Strait. Marine Forecasts (in Chinese), 23(S1): 60–63
|
| [24] |
Sun Jun, Liu Dongyan. 2002. The preliminary notion on nomenclature of common phytoplankton in China Seas waters. Oceanologia et Limnologia Sinica (in Chinese), 33(3): 271–286
|
| [25] |
Sun Jun, Liu Dongyan. 2005. Net phytoplankton community of the Bohai Sea in the autumn of 2000. Haiyang Xuebao (in Chinese), 27(3): 124–132
|
| [26] |
Sun Jun, Liu Dongyan, Qian Shuben. 2002. A quantative research and analysis method for marine phytoplankton: an introduction to Utermöhl method and its modification. Journal of Oceanography of Huanghai & Bohai Seas (in Chinese), 20(2): 105–112
|
| [27] |
Sun Jun, Liu Dongyan, Wei Hao, et al. 2001. Fuzzy evaluation on the marine phytoplankton diversity of Ryükyü-Guntö and its adjacent waters. Oceanologia et Limnologia Sinica (in Chinese), 32(4): 445–453
|
| [28] |
Sun Jun, Song Shuqun, Le Fengfeng, et al. 2007. Phytoplankton in northern South China Sea in the winter of 2004. Haiyang Xuebao (in Chinese), 29(5): 132–145
|
| [29] |
Tan Jiyang, Huang Liangmin, Tan Yehui, et al. 2013. The influences of water-mass on phytoplankton community structure in the Luzon Strait. Haiyang Xuebao (in Chinese), 35(6): 178–189
|
| [30] |
Tang Senming, Cai Rongshuo, Guo Haixia, et al. 2017. Response of phytoplankton in offshore China seas to climate change. Journal of Applied Oceanography (in Chinese), 36(4): 455–465
|
| [31] |
Vila M, Masó M. 2005. Phytoplankton functional groups and harmful algae species in anthropogenically impacted waters of the NW Mediterranean Sea. Scientia Marina, 69(1): 31–45. doi: 10.3989/scimar.2005.69n131
|
| [32] |
Wang Zongling, Li Ruixiang, Zhu Mingyuan, et al. 2006. Study on population growth processes and interspecific competition of Prorocentrum donghaiense and Skeletonema costatum in semi-continuous dilution experiments. Advances in Marine Science (in Chinese), 24(4): 495–503
|
| [33] |
Xie Shangping, Xie Qiang, Wang Dongxiao, et al. 2003. Summer upwelling in the South China Sea and its role in regional climate variations. Journal of Geophysical Research, 108(C8): 3261. doi: 10.1029/2003JC001867
|
| [34] |
Xue Bing, Sun Jun, Li Tingting. 2016. Phytoplankton community structure of northern South China Sea in summer of 2014. Haiyang Xuebao (in Chinese), 38(4): 54–65
|
| [35] |
Yang Dezhou, Yin Baoshu, Liu Zhiliang, et al. 2011. Numerical study of the ocean circulation on the East China Sea shelf and a Kuroshio bottom branch northeast of Taiwan in summer. Journal of Geophysical Research: Oceans, 116(C5): C05015
|
| [36] |
Yin Kedong, Qian Peiyuan, Wu M C S, et al. 2001. Shift from P to N limitation of phytoplankton growth across the Pearl River estuarine plume during summer. Marine Ecology Progress Series, 221: 17–28. doi: 10.3354/meps221017
|
| [37] |
Zhou Hui, Nan Feng, Shi Maochong, et al. 2009. Characteristics of water exchange in the Luzon Strait during September 2006. Chinese Journal of Oceanology and Limnology, 27(3): 650–657. doi: 10.1007/s00343-009-9175-2
|
| [38] |
Zhu Genhai, Ning Xiuren, Cai Yuming, et al. 2003. Studies on species composition and abundance distribution of phytoplankton in the South China Sea. Haiyang Xuebao (in Chinese), 25(S2): 8–23
|
Figures(8) / Tables(4)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: