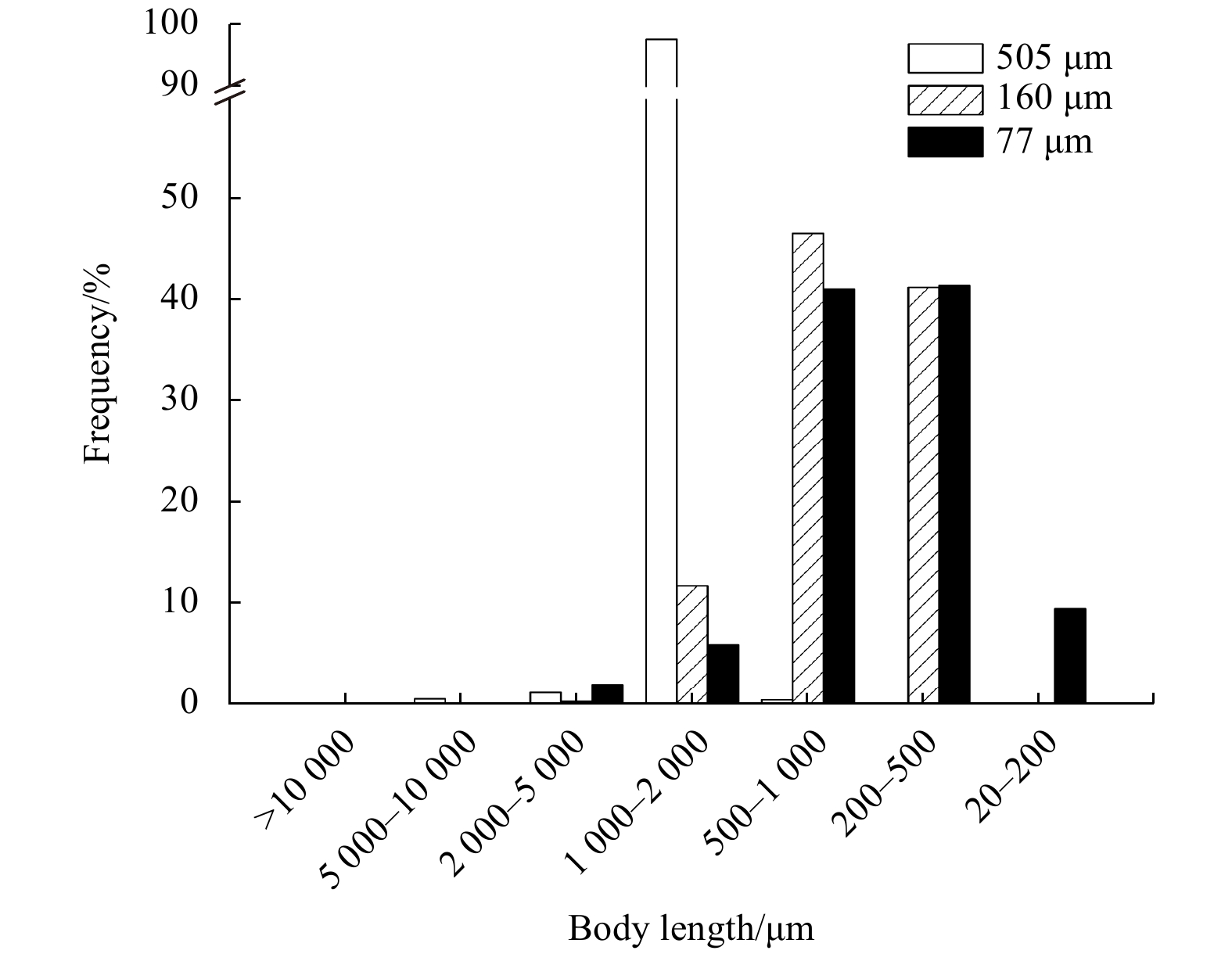
| Citation: | Qianwen Shao, Yifeng Zhu, Meixia Dai, Xia Lin, Chengxu Zhou, Xiaojun Yan. Zooplankton community size-structure change and mesh size selection under the thermal stress caused by a power plant in a semi-enclosed bay[J]. Acta Oceanologica Sinica, 2020, 39(8): 62-70. doi: 10.1007/s13131-020-1634-9

|

| [1] |
Alden R W. 1979. Effects of a thermal discharge on the mortality of copepods in a subtropical estuary. Environmental Pollution, 20(1): 3–19. doi: 10.1016/0013-9327(79)90049-1
|
| [2] |
Antacli J C, Hernández D, Sabatini M E. 2010. Estimating copepods’ abundance with paired nets: Implications of mesh size for population studies. Journal of Sea Research, 63(1): 71–77. doi: 10.1016/j.seares.2009.09.004
|
| [3] |
Bamber R N. 1995. The influence of rising background temperature on the effects of marine thermal effluents. Journal of Thermal Biology, 20(1–2): 105–110. doi: 10.1016/0306-4565(94)00038-K
|
| [4] |
Bernhard M, Möller F, Nassogne A, et al. 1973. Influence of pore size of plankton nets and towing speed on the sampling performance of two high-speed samplers (Delfino I and II) and its consequences for the assessment of plankton populations. Marine Biology, 20(2): 109–136. doi: 10.1007/BF00351450
|
| [5] |
Chew L L, Chong V C, Wong R C S, et al. 2015. Three decades of sea water abstraction by Kapar power plant (Malaysia): What impacts on tropical zooplankton community?. Marine Pollution Bulletin, 101(1): 69–84. doi: 10.1016/j.marpolbul.2015.11.022
|
| [6] |
Clarke K R, Gorley R N, Somerfield P J, et al. 2014. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. Plymouth, UK: Primer-E Ltd, 256
|
| [7] |
Cushing D H, Humphrey G F, Banse K, et al. 1958. Report of the committee on terms and equivalents. Rapp P-V. Réun Cons Int Explor Scient Mer Méditerr, 144: 15–16
|
| [8] |
Davies R M, Jensen L D. 1975. Zooplankton entrainment at three mid-Atlantic power plants. Journal-Water Pollution Control Federation, 47(8): 2130–2142
|
| [9] |
Du Xiuning, Wang Yunlong, Chen Tao, et al. 2013. Seasonal variation of zooplankton copepod community in Xiangshan Bay of East China. Chinese Journal of Ecology (in Chinese), 32(10): 2756–2763
|
| [10] |
Du Xiuning, Wang Yunlong. 2014. Inter-annual and seasonal changes of zooplankton biomass and species in the specific region of the Xiangshan Bay. Marine Science Bulletin (in Chinese), 33(3): 293–298
|
| [11] |
Du Ping, Xu Xiaoqun, Liu Jingjing, et al. 2015. Spatial heterogeneity of macro-and meso-zooplankton in Xiangshan Bay in spring and summer. Acta Ecologica Sinica (in Chinese), 35(7): 2308–2321. doi: 10.5846/stxb201306091487
|
| [12] |
Du Ping, Xu Xiaoqun, Xu Xudan, et al. 2017. Effects of three different aquaculture activities on zooplankton community in Xiangshan Bay. Journal of Fisheries of China (in Chinese), 41(11): 1719–1733
|
| [13] |
Dussart B M. 1965. Les différentes catégories de plancton. Hydrobiologia, 26(1): 72–74. doi: 10.1007/BF00142255
|
| [14] |
Evans M S, Warren G J, Page D I. 1986. The effects of power plant passage on zooplankton mortalities: Eight years of study at the Donald C. Cook nuclear plant. Water Research, 20(6): 725–734. doi: 10.1016/0043-1354(86)90096-5
|
| [15] |
Gallienne C P, Robins D B. 2001. Is Oithona the most important copepod in the world’s oceans?. Journal of Plankton Research, 23(12): 1421–1432. doi: 10.1093/plankt/23.12.1421
|
| [16] |
Hwang J S, Kumar R, Dahms H U, et al. 2007. Mesh size affects abundance estimates of Oithona spp. (Copepoda, Cyclopoida). Crustaceana, 80(7): 827–837. doi: 10.1163/156854007781363169
|
| [17] |
Jiang Zhibing, Zeng Jiangning, Chen Quanzhen, et al. 2009. Potential impact of rising seawater temperature on copepods due to coastal power plants in subtropical areas. Journal of Experimental Marine Biology and Ecology, 368(2): 196–201. doi: 10.1016/j.jembe.2008.10.016
|
| [18] |
Jiang Zhibing, Chen Quanzhen, Zeng Jiangning, et al. 2012. Phytoplankton community distribution in relation to environmental parameters in three aquaculture systems in a Chinese subtropical eutrophic bay. Marine Ecology Progress Series, 446: 73–89. doi: 10.3354/meps09499
|
| [19] |
Jones K J, Gowen R J. 1990. Influence of stratification and irradiance regime on summer phytoplankton composition in coastal and shelf seas of the British Isles. Estuarine, Coastal and Shelf Science, 30(6): 557–567. doi: 10.1016/0272-7714(90)90092-6
|
| [20] |
Laws E A, Falkowski P G, Smith Jr W O, et al. 2000. Temperature effects on export production in the open ocean. Global Biogeochemical Cycles, 14(4): 1231–1246. doi: 10.1029/1999GB001229
|
| [21] |
Liu Zilin, Cai Yuming, Ning Xiuren. 1998. The distribution of chlorophyll a and primary productivity in the middle and west of Xiangshan Bay. Donghai Marine Science (in Chinese), 16(3): 18–24
|
| [22] |
Liu Zhensheng, Wang Chunsheng, Yang Junyi, et al. 2004. Distribution of zooplankton in the Xiangshangang Bay in winter. Donghai Marine Science (in Chinese), 22(1): 34–42
|
| [23] |
Lloyd P, Plagányi É E, Weeks S J, et al. 2011. Ocean warming alters species abundance patterns and increases species diversity in an African sub-tropical reef-fish community. Fisheries Oceanography, 21(1): 78–94. doi: 10.1111/j.1365-2419.2011.00610.x
|
| [24] |
Mäkinen K, Vuorinen I, Hänninen J. 2017. Climate-induced hydrography change favours small-bodied zooplankton in a coastal ecosystem. Hydrobiologia, 792(1): 83–96. doi: 10.1007/s10750-016-3046-6
|
| [25] |
Miao Qingsheng, Zhou Liangming, Deng Zhaoqing. 2010. Numerical simulation and in-situ measurement for heat discharge from Xiangshangang power plant Into Sea. Coastal Engineering, 29(4): 1–11
|
| [26] |
Nielsen T G, Sabatini M. 1996. Role of cyclopoid copepods Oithona spp. in North Sea plankton communities. Marine Ecology Progress Series, 139: 79–93. doi: 10.3354/meps139079
|
| [27] |
Pansera M, Granata A, Guglielmo L, et al. 2014. How does mesh-size selection reshape the description of zooplankton community structure in coastal lakes?. Estuarine, Coastal and Shelf Science, 151: 221–235. doi: 10.1016/j.ecss.2014.10.015
|
| [28] |
Porter K G. 1973. Selective grazing and differential digestion of algae by zooplankton. Nature, 244(5412): 179–180. doi: 10.1038/244179a0
|
| [29] |
Riccardi N. 2010. Selectivity of plankton nets over mesozooplankton taxa: implications for abundance, biomass and diversity estimation. Journal of Limnology, 69(2): 287–296. doi: 10.4081/jlimnol.2010.287
|
| [30] |
Rice E, Dam H G, Stewart G. 2015. Impact of climate change on estuarine zooplankton: surface water warming in long island sound is associated with changes in copepod size and community structure. Estuaries and Coasts, 38(1): 13–23. doi: 10.1007/s12237-014-9770-0
|
| [31] |
Stabeno P J, Kachel N B, Moore S E, et al. 2012. Comparison of warm and cold years on the southeastern Bering Sea shelf and some implications for the ecosystem. Deep Sea Research Part II: Topical Studies in Oceanography, 65–70: 31–45. doi: 10.1016/j.dsr2.2012.02.020
|
| [32] |
Şundri M I, Gomoiu M T. 2009. Qualitative and quantitative structure of zooplankton asociations in the Danube thermal discharge area of Nuclear Power Plant Cernavoda. Geoecomarina, 15: 123–130
|
| [33] |
Tseng L C, Dahms H U, Hung J J, et al. 2011. Can different mesh sizes affect the results of copepod community studies?. Journal of Experimental Marine Biology and Ecology, 398(1–2): 47–55. doi: 10.1016/j.jembe.2010.12.007
|
| [34] |
Tunowski J. 2009. Changes in zooplankton abundance and community structure in the cooling channel system of the Konin and Pątnów power plants. Fisheries & Aquatic Life, 17(4): 279–289. doi: 10.2478/v10086-009-0020-1
|
| [35] |
Turner J T. 2004. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zoological Studies, 43(2): 255–266
|
| [36] |
Vannucci M. 1968. Loss of organisms through the meshes. In: Tranter D J, ed. Zooplankton Sampling. UNESCO Monographs on Oceanographic Methodology. Paris: UNESCO Press, 77–86
|
| [37] |
Wang Chunsheng, Liu Zhensheng, He Dehuai. 2003. Seasonal dynamics of zooplankton biomass and abundance in Xiangshan Bay. Journal of Fisheries of China (in Chinese), 27(6): 595–599
|
| [38] |
Wang Xiaobo, Qiu Wusheng, Qin Mingli, et al. 2009. Studies on ecological community distribution of zooplankton in Xiangshan Bay. Marine Environmental Science (in Chinese), 28(1): 62–64
|
| [39] |
Wang Yangcai, Wu Xiongfei, Shi Huixiong, et al. 2011. Study on zooplankton communities near coastal power plants in Xiangshan Bay. Journal of Ningbo University (Natural Science & Engineering Edition) (in Chinese), 24(3): 5–10
|
| [40] |
Xu Zhaoli, Chen Yaqu. 1989. Aggregated intensity of dominant species of zooplankton in autumn in the East China Sea and Yellow Sea. Journal of Ecology (in Chinese), 8(4): 13–15
|
| [41] |
Yvon-Durocher G, Montoya J M, Trimmer M, et al. 2011. Warming alters the size spectrum and shifts the distribution of biomass in freshwater ecosystems. Global Change Biology, 17(4): 1681–1694. doi: 10.1111/j.1365-2486.2010.02321.x
|
| [42] |
Zheng Zhong, Li Song, Li Shaojing, et al. 1982. Pelagic Copepoda in China Seas (in Chinese). Shanghai: Shanghai Science and Technology Press, 1–162
|
| [43] |
Zheng Zhong, Li Shaojing, Xu Zhenzu, et al. 1984. Marine Planktology (in Chinese). Beijing: China Ocean Press, 1–653
|

Figures(5) / Tables(6)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: