School of Environment, Harbin Institute of Technology, Harbin 150090, China
2.
Key Laboratory of Marine Genetic Resources, Third Institute of Oceanography, Ministry of Natural Resources, Xiamen 361005, China
3.
State Key Laboratory Breeding Base of Marine Genetic Resources, Xiamen 361005, China
4.
Fujian Key Laboratory of Marine Genetic Resources, Xiamen 361005, China
5.
Sourthern Marine Science and Engineering Guangdong Laboratory (Zhuhai), Zhuhai 519080, China
Funds:
The COMRA Program under contract No. DY135-B2-01; the Xiamen Ocean Economic Innovation and Development Demonstration Project under contract No. 16PZP001SF16; the National Infrastructure of Natural Resources for Science and Technology Program of China under contract No. NIMR-2017-9.
Nitrification is a key step in the global nitrogen cycle. Compared with autotrophic nitrification, heterotrophic nitrification remains poorly understood. In this study, Halomonas venusta MA-ZP17-13, isolated from seawater in shrimp aquaculture (Penaeus vannamei), could simultaneously undertake nitrification and denitrification. With the initial ammonium concentration at 100 mg/L, the maximum ammonium-nitrogen removal rate reached 98.7% under the optimal conditions including C/N concentration ratio at 5.95, pH at 8.93, and NaCl at 2.33%. The corresponding average removal rate was 1.37 mg/(L·h) (according to nitrogen) in 3 d at 11.2°C. By whole genome sequencing and analysis, nitrification- and denitrification-related genes were identified, including ammonia monooxygenase, nitrate reductase, nitrite reductase, nitric oxide dioxygenase and nitric oxide synthase; while no gene encoding hydroxylamine oxidase was identified, it implied the existence of a novel nitrification pathway from hydroxylamine to nitrate. These results indicate heterotrophic bacterium H. venusta MA-ZP17-13 can undertake simultaneous nitrification and denitrification at low temperature and has potential for ${\rm {NH}}_4^+ $-N/NH3-N removal in marine aquaculture systems.
Nitrogen (N) compounds are indispensable in biological metabolic processes and play vital roles in nutrient cycling. Inorganic ammonium (${\rm {NH}}_4^+ $) is generally considered an undesirable product because it can cause water quality problems and has a pungent smell (He et al., 2017). Water quality is crucial to the aquaculture industry, and the concentration of ${\rm {NH}}_4^+ $-N/NH3-N is an important indicator of water quality in aquaculture. Many N-containing organic compounds can be excreted under high feed loading rate of aquaculture organisms, which can generate a large number of harmful N-containing substances such as NH3 and nitrite (${\rm {NO}}_2^- $), causing acute poisoning of aquaculture organisms (Shan and Obbard, 2001; Lin and Chen, 2003). High concentrations of NH3 in aquaculture water will affect the cell membrane and enzymatic activity of aquaculture organisms, and even destroy the excretory system and osmotic balance in serious cases (Atanassov et al., 1994). NH3-N in water is composed of two forms: ionized ammonia (or ammonium: ${\rm {NH}}_4^+ $-N) and unionized ammonia (NH3-N) (Armstrong et al., 1978; Thurston et al., 1981). Compared with ${\rm {NH}}_4^+ $-N, NH3-N is more toxic, because the neutral charge and small size of the molecule that can easily pass through cell membrane (Smart, 1978). It was reported that the 48 h lethal concentration (LC50) values for NH3-N to Litopenaeus vannamei were 2.09 mg/L to 2.26 mg/L in shrimp aquaculture (Schuler et al., 2010). Therefore, controlling the NH3-N concentration in aquaculture water is particularly important.
Generally, the treatment of ${\rm {NH}}_4^+ $-N/NH3-N is mainly carried out by physical and biological methods in aquaculture. The main physical method is to regularly sprinkle zeolite powder or Maifan stone into the water (Wei et al., 2010; Kan et al., 2011), but this approach is a palliative rather than a cure; the most common and cost-effective method for ${\rm {NH}}_4^+ $-N/NH3-N removal is biological treatment, which involves a combined process of autotrophic nitrification and heterotrophic denitrification (Kim et al., 2005; Chiu et al., 2007; Zhu et al., 2008; Zhao et al., 2010a; Huang et al., 2013). However, this conventional method tends to be time-consuming due to the low rate of nitrification and the separation of aerobic and anoxic phases (Zhu et al., 2008; Huang et al., 2013).
To obtain bacteria that are capable of effective ${\rm {NH}}_4^+ $-N/NH3-N removal at low temperature, bacteria were enriched and isolated from surface seawater of a higher-place pond of Penaeus vannamei. Among the obtained isolates, Halomonas venusta MA-ZP17-13 showed high efficiency in removing ${\rm {NH}}_4^+ $-N/NH3-N with no accumulation of ${\rm {NO}}_2^- $-N at 10°C. Moreover, it was also capable for heterotrophic nitrification and aerobic denitrification. The aims of this study are (1) to obtain bacteria that are capable for effective ${\rm {NH}}_4^+ $-N/NH3-N removal at low temperature; (2) to study the characteristics of ${\rm {NH}}_4^+ $-N/NH3-N removal of starin MA-ZP17-13 at low temperature. Studies indicate strain MA-ZP17-13 has a great potential in ${\rm {NH}}_4^+ $-N/NH3-N removal from aquaculture water at low temperature.
2.
Materials and methods
2.1
Sampling and strain isolation
Surface seawater of higher-place P. vannamei ponds was sampled in June 2018. The aquaculture seawater sample (24.02°N, 117.83°E) was obtained from an aquaculture farm in Zhangpu County, Zhangzhou City, Fujian Province, China. The seawater was diluted and spread on heterotrophic nitrification medium (HNM) (Xu et al., 2017). After 4 d of aerobic incubation at 28°C, single colonies were picked. Purity was confirmed by the uniformity of cell morphology after repeated streaking. During bacterial screening process, a strain named MA-ZP17-13 was isolated along with many other bacterial isolates. For morphological and biochemical characterization, strain MA-ZP17-13 was cultivated on marine agar 2216 medium. For long-time storage, the strain was suspended in 20.0% glycerol solution at −80°C and deposited in Marine Culture Collection of China under accession number MCCC 1A14584.
2.2
Culture media
Marine agar 2216 medium (BD Difco) (Li et al., 2019) contained (per L): 5.0 g peptone, 1.0 g yeast extract, 0.1 g FeC6H5O7, 19.45 g NaCl, 8.8 g MgCl2, 3.24 g Na2SO4, 1.8 g CaCl2, 0.55 g KCl, 0.16 g NaHCO3, 0.08 g KBr, 34.0 mg SrCl2, 22.0 mg H3BO3, 4.0 mg Na2SiO3, 2.4 mg NaF, 1.6 mg NH4NO3, 8.0 mg Na2HPO4.
HNM (pH=7.2) (Xu et al., 2017) contained (per L): 0.24 g (NH4)2SO4, 1.12 g C4H4Na2O4·6H2O, 0.05 g MgSO4·7H2O, 0.5 g KH2PO4, 2.5 g K2HPO4, 30 g NaCl, 0.05 g MnSO4, 0.05 g FeSO4 and 1 mL trace element solution. HNM medium (inorganic medium) was used to determine the ${\rm{NH}}_4^+$-N removal ability of the isolate strain.
Denitrification medium (DM; pH = 7.2) (Xu et al., 2017) contained (per L) : 1.08 g KNO3, 8.43 g C4H4Na2O4·6H2O, 7.9 g Na2HPO4·7H2O, 1.5 g KH2PO4, 0.1 g MgSO4·7H2O, 30 g NaCl, 1 mL trace element solution. The isolate was cultivated in DM medium to test the ability of denitrification.
Trace element solution contained (per L): 2 mg CaCl2, 50 mg FeCl3·6H2O, 0.5 mg CuSO4, 0.5 mg MnCl2·4H2O and 10 mg ZnSO4·7H2O. Cultures were incubated at 10°C and spun at 150 r/min, unless noted otherwise.
2.3
Estimation the N removal and conversion capacity of strain MA-ZP17-13
The strain MA-ZP17-13 was activated on marine agar 2216 plates, and inoculated into 100 mL marine broth 2216 medium and cultured at 150 r/min and 10°C for 24 h. The culture of 2 mL was centrifuged to remove the medium and washed twice with sterilized seawater, then inoculated into 100 mL HNM medium for ammonia removal tests. After incubated at 10°C and 150 r/min for 15 h under aerobic conditions, 5 mL liquid cultures were sampled serially at intervals of several hours to detect the concentrations of ${\rm {NH}}_4^+ $-N, nitrate (${\rm {NO}}_3^- $-N) and nitrite (${\rm {NO}}_2^- $-N). All experiments were conducted in triplicate.
The ${\rm {NH}}_4^+ $-N removal efficiency was calculated by the equation: Rv=[(T1−T2)/T1]×100.0%, where Rv, T1 and T2 represented ${\rm {NH}}_4^+ $-N removal efficiency, and the initial and final ${\rm {NH}}_4^+ $-N concentration, respectively. The heterotrophic nitrification rate and the ${\rm {NO}}_3^- $-N removal rate were calculated by linear fitting of the concentration changes of ${\rm {NO}}_2^- $-N, ${\rm {NO}}_3^- $-N and ${\rm {NO}}_3^- $-N, respectively. The heterotrophic nitrification rate was measured with 0.01% (v/v) acetylene inhibition, according to acetylene could inhibit the denitrification process and the growth of autotrophic bacteria (Ryden et al., 1987; McCarty, 1999; Lu and Jia, 2013; Zhang et al., 2014).
To detect bacterial ammonia assimilation, strain MA-ZP17-13 was cultured in HNM at 150 r/min and 10°C for 72 h in 100 mL flask. After centrifugation and freeze-drying, the intracellular N content was detected by elemental analyzer EL-III (Vario EL-III). The intracellular N content was calculated by the formula as follows: R=R1×M/V, where R (mg/L), R1 (%), M (mg) and V (L) represented ${\rm {NH}}_4^+ $-N/NH3-N concentration, the percentage of nitrogen content, dry weight, and medium volume, respectively.
2.4
Quantification methods of ${\boldsymbol {NH}}_{\text{4}}^+ $, ${\boldsymbol {NO}_{\text{2}}^-} $ and ${\boldsymbol {NO}}_{\text{3}}^-$
The concentrations of ${\rm {NH}}_4^+ $-N, ${\rm {NO}}_2^- $-N and ${\rm {NO}}_3^- $-N were detected using the supernatant after samples centrifuged at 8 000 r/min for 5 min. The concentrations of ${\rm {NH}}_4^+ $-N, ${\rm {NO}}_3^- $-N and ${\rm {NO}}_2^- $-N were determined according to standard methods (APHA, 2005). The ${\rm {NH}}_4^+ $-N concentration was analyzed by the method of Nessler’s reagent spectrophotometry. The ${\rm {NO}}_3^- $-N concentration was calculated by subtracting two times the background absorbance value at 275 nm from the absorbance value at 220 nm. ${\rm {NO}}_2^- $-N was determined at wavelengths of 540 nm after adding 1 mL chromogenic reagent including 0.1 mL phosphoric acid, 0.002 g N-(1-naphthyl)-1,2-diaminoethane dihydrochloride and 0.04 g sulfanilamide (APHA, 2005). In addition, N2O and N2 were detected on a gas chromatography equipped with a thermal conductivity detector and electron capture detector, respectively (Zhao et al., 2012).
2.5
Genomic DNA preparation
For genome sequencing, strain MA-ZP17-13 was grown aerobically to mid logarithmic phase at 10°C on marine agar 2216 medium. Genomic DNA was isolated from the cell pellets using the ChargeSwitch® gDNA Mini Bacteria Kit (Life Technologies) according to the manufacturer’s instructions. Purified genomic DNA was quantified by TBS-380 fluorometer (Turner BioSystems, CA). High quality DNA (OD260/280 is 1.8−2.0, >10 μg) was used to do further research.
2.6
Genome sequencing and analysis
The genome of strain MA-ZP17-13 was sequenced using a combination of PacBio RS and Illumina sequencing platforms. The Illumina data was used to evaluate the complexity of the genome. These data were tried to be assembled using Velvet assembler (v1.2.09) with a k-mer length of 17 (Zerbino, 2010). The complete genome sequence was assembled using both the PacBio reads and Illumina reads. The assembly was produced firstly using a hybrid de novo assembly solution modified by Koren (Koren et al., 2012), in which a de-Bruijn based assembly algorithm and a CLR (continuous long reads) reads correction algorithm were integrated in “PacBioToCA with Celera Assembler” pipeline (Chin et al., 2013). The last circular step was checked and finished manually. The final assembly generated a circular genome sequence with no gap. The complete genome sequence was submitted to GenBank under accession No. CP034367.
Identification of predicted coding sequences (CDS), also called open reading frames (ORFs), was performed using Glimmer version 3.02. ORFs with less than 300 bp were discarded. Then remaining ORFs were queried against the non-redundant (NR) protein database in the National Center for Biotechnology Information (NCBI), SwissProt (http://uniprot.org), Kyoto Encyclopedia of Genes and Genomes (KEGG, http://www.genome.jp/kegg/), Gene Ontology (GO, http://geneontology.org) and Phylogenetic Classification of Proteins Encoded in Complete Genomes (COG, http://www.ncbi.nlm.nih.gov/COG) databases for functional annotation. In addition, plasmids were identified using the GeneMarkS (v4.30), tRNAs were identified using the tRNAscan-SE (v1.3.1) and rRNA were determined using Barrnap (v0.4.2). Circos software was then used to generate a circularized map of the chromosome, setting the calculation window at 2 000 bp with steps of 500 bp (Krzywinski et al., 2009).
Gene prediction and annotation were performed using the NCBI prokaryotic genome annotation pipeline (Tatusova et al., 2016) and six large databases (NR, Swiss-Prot, Pfam, COG, GO, and KEGG). The functional annotation of predicted ORFs was used to search the KEGG and COG databases by RPS-BLAST.
2.7
Bacterial identification based on whole genomic sequences
The average nucleotide identity (ANI) between strains MA-ZP17-13 and H. venusta DSM 4743T was calculated with EZGenome using the algorithm of Goris et al. (2007). DNA-DNA hybridization (DDH) estimate values between two strains were analyzed using the genome-to-genome distance calculator (GGDC2.0) (Auch et al., 2010a, b; Meier-Kolthoff, et al., 2013).
2.8
Optimization of cultivation parameters using response surface methodology
The critical factors affecting ${\rm {NH}}_4^+ $-N removal efficiency, including C/N concentration ratio, pH, salinity and temperature, were screened by single-factor experiment.
Based on the preliminary results, the appropriate range of independent variables including C/N concentration ratio (A), pH (B), salinity (C) and temperature (D) were determined. The response surface methodology is an important math-statistics technique for designing, modeling and analysis of problems where a response of interest is affected by several different parameters (Qu et al., 2017). Design expert 8.0.5b software was used for the design of the experimental run (Myers et al., 2016). Box-Behnken design (BBD) was applied to generate the design matrix of four factors at three levels (Table 1). Twenty-nine BBD trials were conducted to provide direction for further optimization studies (Table 2). Analysis of variance (ANOVA) was used to check the validity of regression model and determine the quadratic effect of machine parameters on the output response function.
Table
1.
Real and coded variables used to optimize ammonia nitrogen removal in BBD
Statistical model analysis was evaluated to determine the ANOVA and the quadratic models were constituted as 3D contour plots using Design-Expert® 8 software (Myers et al., 2016).
3.
Results and discussion
3.1
Identification of strain MA-ZP17-13
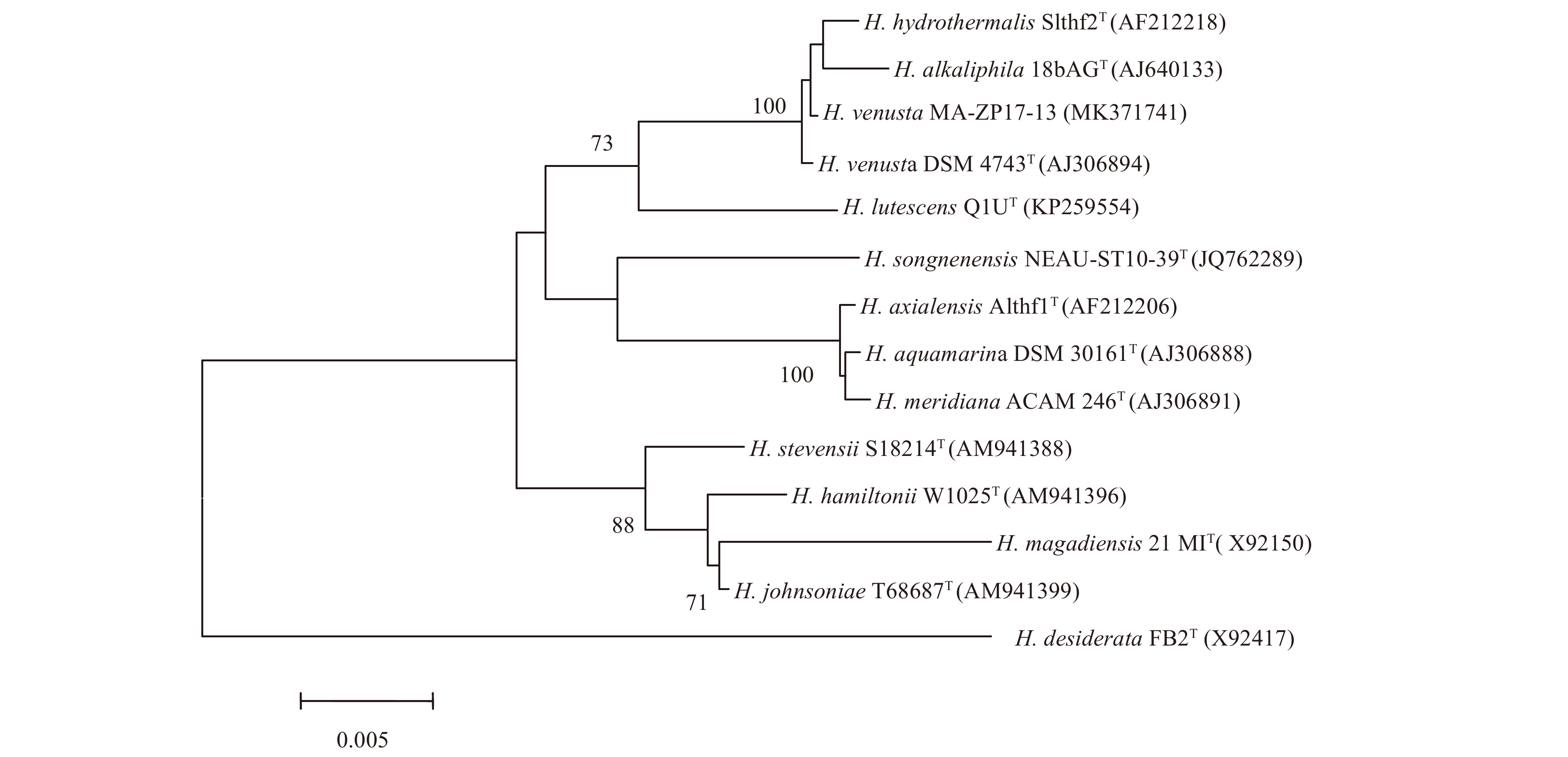
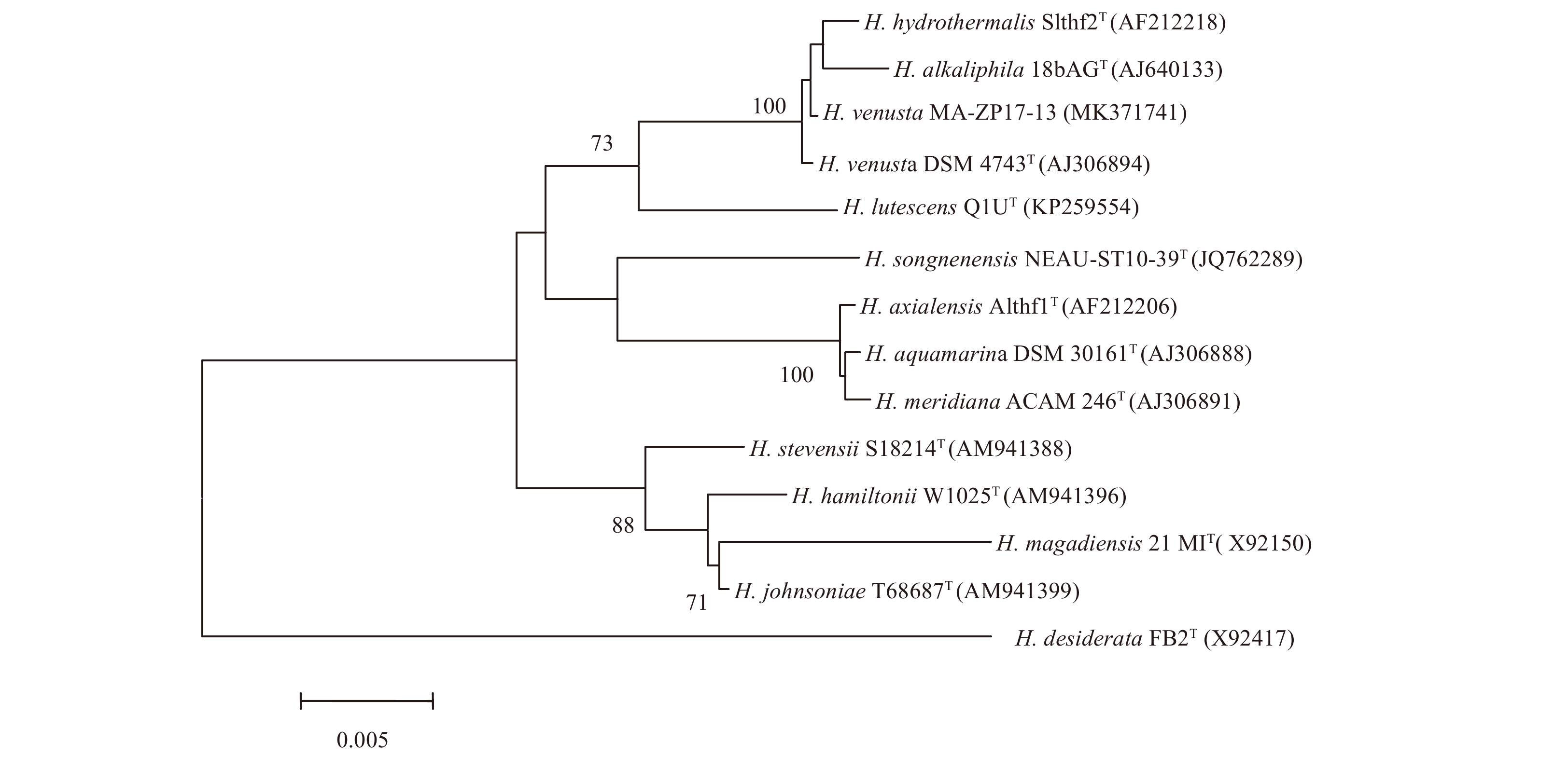
A nearly full-length 16S rRNA gene sequence (1 540 nt) of strain MA-ZP17-13 was determined, which showed the highest sequence similarity to H. venusta DSM 4743T (99.93%), followed by H. hydrothermalis Slthf2T (99.79%) and H. alkaliphila 18bAGT (99.73%). A phylogenetic tree was constructed based on the 16S rRNA gene sequences of the genus Halomonas (Fig. 1), which showed that strain MA-ZP17-13 formed a clade with H. venusta DSM 4743T. The data DNA-DNA hybridization estimate value between strain MA-ZP17-13 and H. venusta DSM 4743T was 90.80%, which was above the standard cut-off value (70%) (Wayne et al., 1987). The ANI value between strain MA-ZP17-13 and H. venusta DSM 4743T was 98.78%, which was above the standard ANI criteria for species identity (95%–96%) (Richter and Rosselló-Móra, 2009). Hence, strain MA-ZP17-13 was identified as H. venusta based on 16S rRNA gene sequence analyses and dDDH value.
Figure
1.
Neighbour-joining tree showing the phylogenetic positions of strain MA-ZP17-13 and other members of the genus Halomonas, based on 16S rRNA gene sequences. The bootstrap values on the branching nodes were calculated on 1 000 replications. The scale bar indicated 0.005 substitutions per nucleotide position.
On marine agar 2216, colonies of strain MA-ZP17-13 are milk white, opaque, convex, regular with entire margin and 1.0–2.0 mm in diameter after 2 d at 28°C. General features of strain MA-ZP17-13 are summarized in Table 3.
Table
3.
General features of strain MA-ZP17-13 and MIGS mandatory information
The strain is positive for catalase and oxidase, reduction of nitrate and denitrification, and is negative for arginine dihydrolase, indole production, D-glucose fermentation, urease, β-galactosidase. API ZYM test strip results indicate that it is positive for alkaline phosphatase, esterase (C4), lipase (C14), leucine aminopeptidase, naphtol-AS-BI-phosphoamidase, valine aminopeptidase, acid phosphatase, α-glucosidase; it is weakly positive for esterase lipase (C8), cystine aminopeptidase; it is negative for N-acetyl-β-glucosaminidase, trypsin, α-chymotrypsin, α-galactosidase, β-galactosidase, β-glucuronidase, β-glucosidase, α-fucosidase and α-mannosidase. The API 20NE test strip shows that strain MA-ZP17-13 can utilize D-glucose, D-mannitol, D-maltose, N-acetyl-glucosamine, potassium gluconate, capric acid, adipic acid, malic acid, trisodium citrate and phenylacetic acid, cannot utilize L-arabinose and D-mannose.
3.2
Determination of the ${\boldsymbol {NH}}_{\text{4}}^+ $-N/NH3-N removal rate and transformation capacity of strain MA-ZP17-13
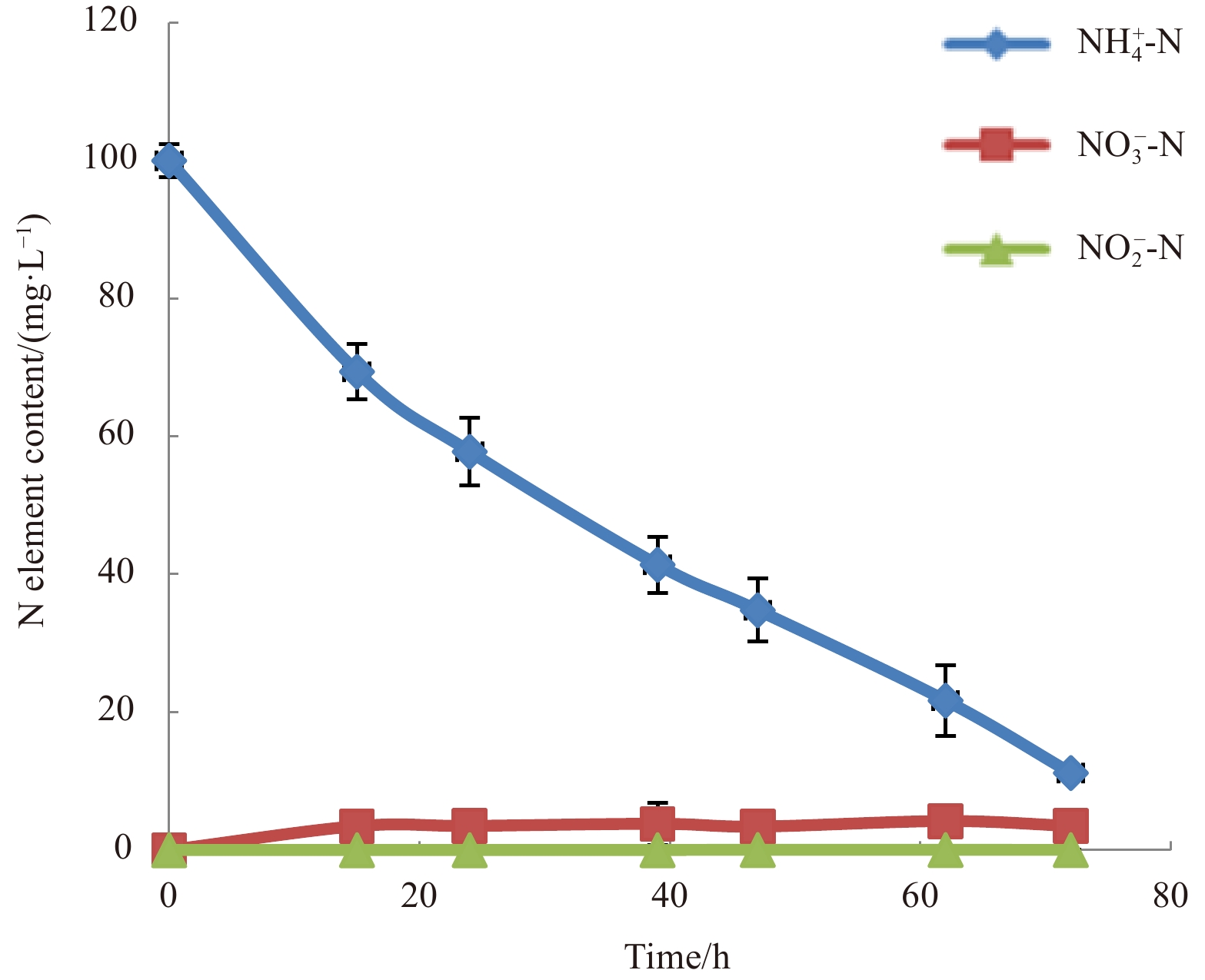
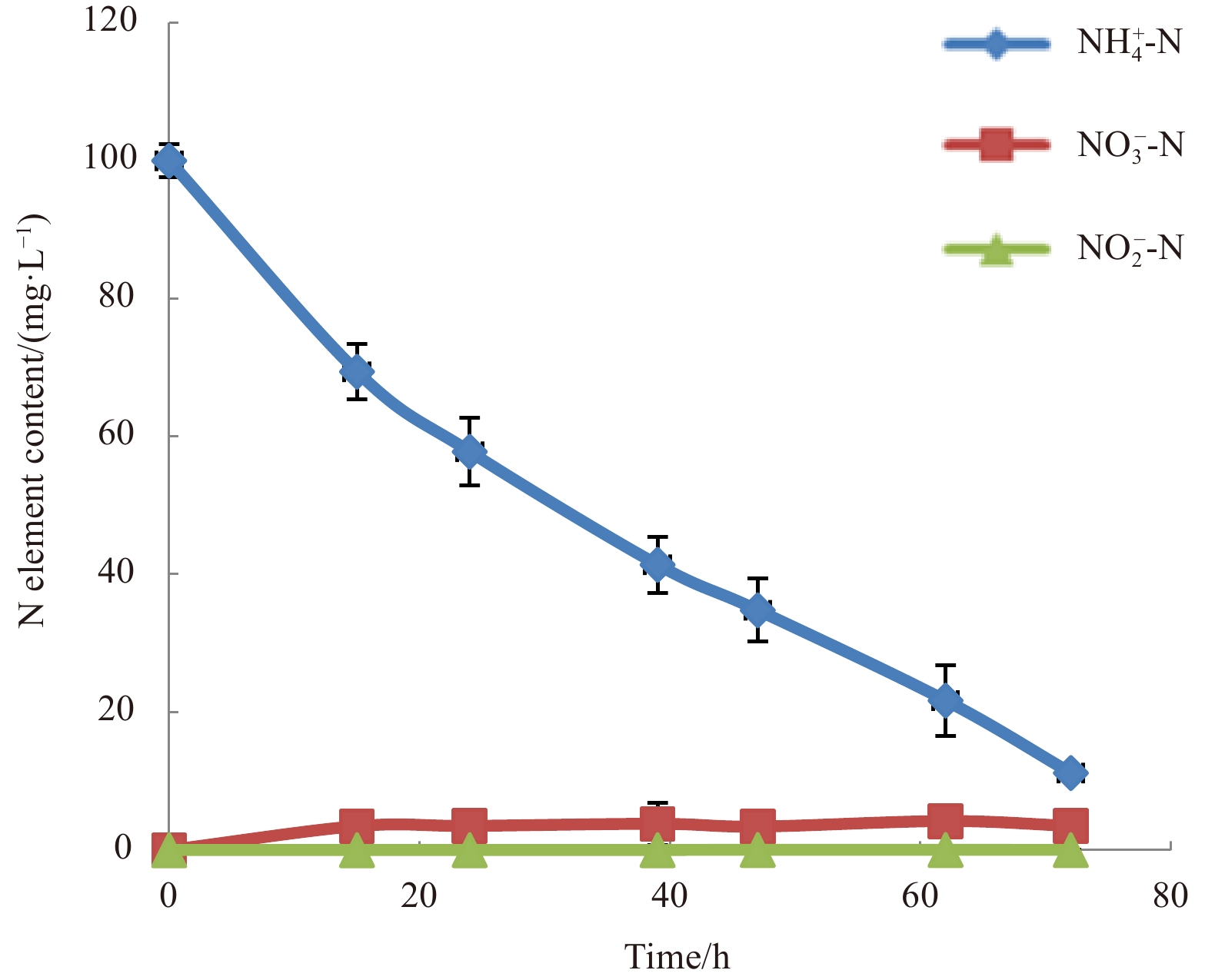
Low temperature can reduce ${\rm {NH}}_4^+ $-N/NH3-N removal rate by slowing down microbial growth (Zhang et al., 2011; Rodriguez-Caballero et al., 2012; Huang et al., 2013). To determine the ${\rm {NH}}_4^+ $-N/NH3-N removal efficiency of strain MA-ZP17-13 at 10°C, 100 mg/L initial concentration of ${\rm {NH}}_4^+ $-N/NH3-N was used as the sole nitrogen source in HNM medium. The ${\rm {NH}}_4^+ $-N/NH3-N concentration showed a significant decrease from 100.0 mg/L to 11.2 mg/L, with about 90% of ${\rm {NH}}_4^+ $-N/NH3-N removal after 72 h of incubation at 10°C, and the average ${\rm {NH}}_4^+ $-N/NH3-N removal rate was approximately 1.23 mg/(L·h) (Fig. 2). With the increase of culture time, ${\rm {NH}}_4^+ $-N/NH3-N could not be removed completely, but it could be decreased to 3 mg/L within 120 h. Microbacterium sp. SFA13 was reported to have the ability of ${\rm {NH}}_4^+ $-N/NH3-N removal, with an average removal rate of 0.11 mg/(L·h) at 5°C (Zhang et al., 2013). Strain MA-ZP17-13 showed a high potential for ${\rm {NH}}_4^+ $-N/NH3-N removal at low temperature. Therefore, this strain has important application potential in sewage treatment of ${\rm {NH}}_4^+ $-N/NH3-N removal under low temperature.
Figure
2.
Ammonium-nitrogen removal rate of strain Halomonas venusta MA-ZP17-13.
The ${\rm {NH}}_4^+ $-N/NH3-N removal in the medium is supposed to be carried out via assimilation and/or nitrification. To detect the amount of ${\rm {NH}}_4^+ $-N/NH3-N assimilation, strain MA-ZP17-13 was cultured in HNM medium at the initial concentration of 100 mg/L ${\rm {NH}}_4^+ $-N, at 150 r/min and 10°C for 72 h. After centrifuging and freeze-drying, the average dry weight of three parallel samples was 48.4 mg. The average percentage of nitrogen concentration is 10.4%. After calculation by the equation R=R1×M/V, the ${\rm {NH}}_4^+ $-N/NH3-N assimilated by strain MA-ZP17-13 was 50.3 mg/L on average.
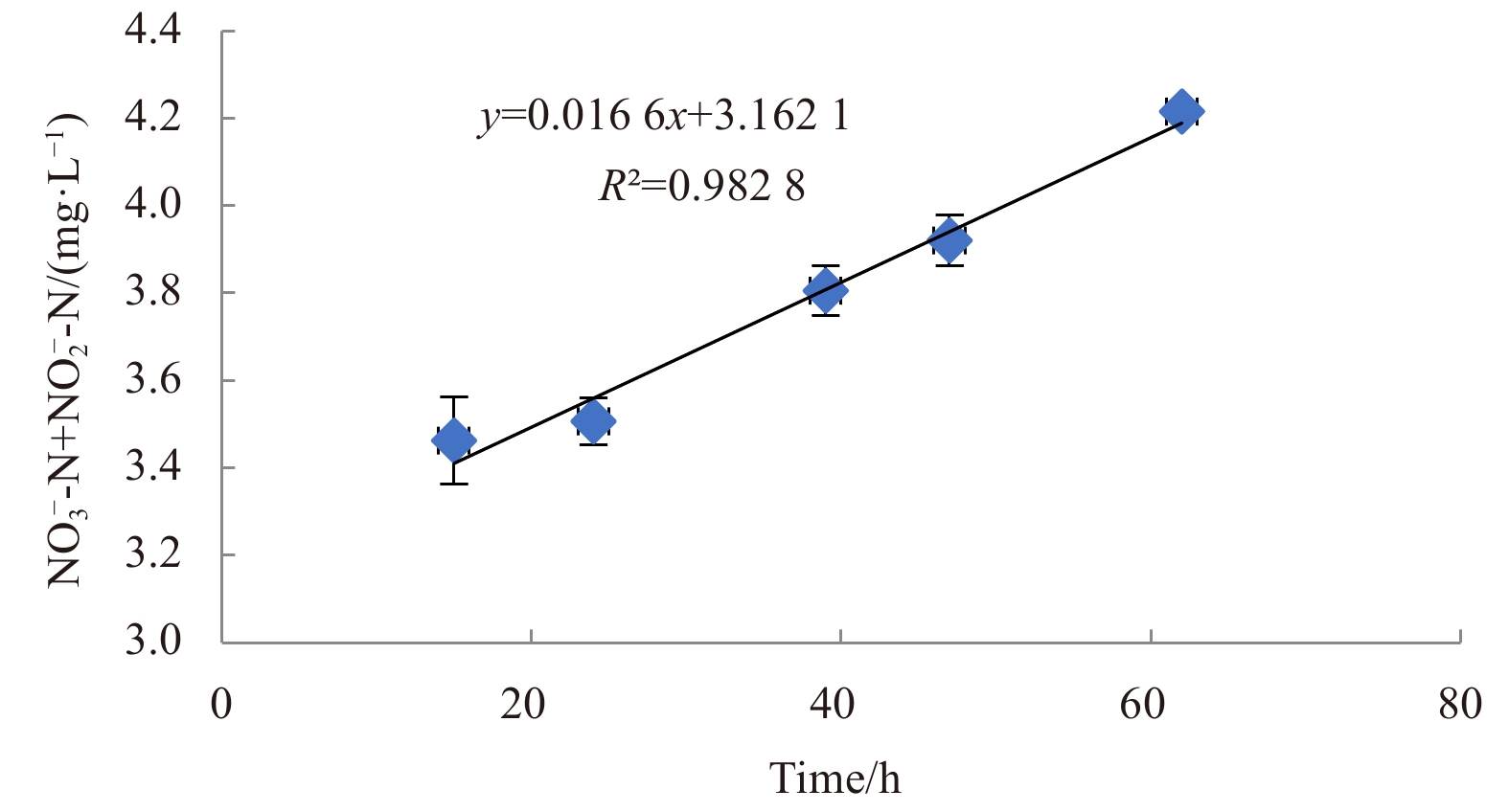
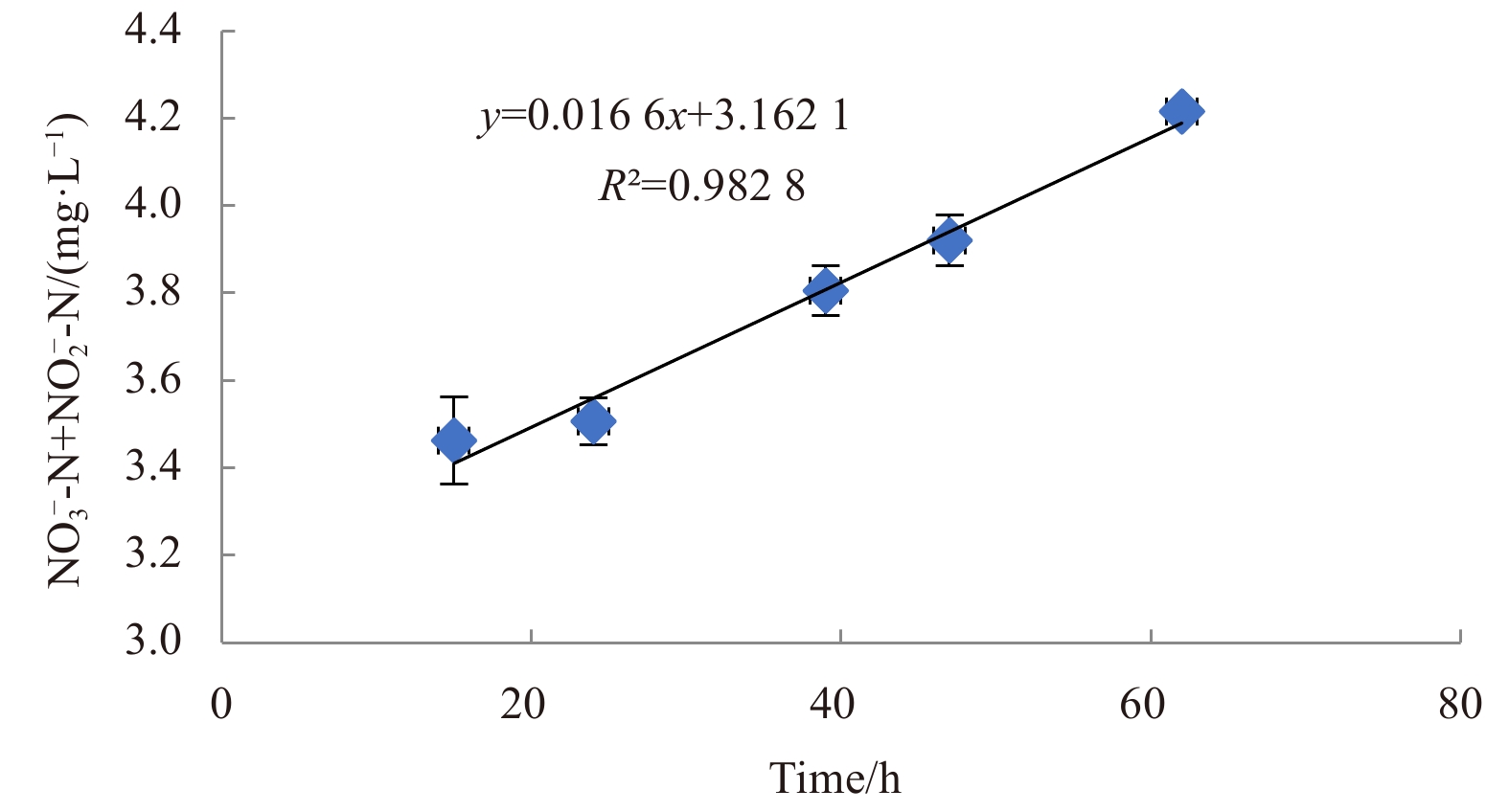
To determine nitrification efficiency of this bacterium at low temperature, ammonia was used as the only N source in HNM medium under aerobic conditions at 10°C, with an initial concentration of 100 mg/L ${\rm {NH}}_4^+ $-N/NH3-N. The slope of the linear line of best fit was equal to the heterotrophic nitrification rate (Fig. 3). The heterotrophic nitrification rate was 0.016 6 mg/(L·h). The average ${\rm {NH}}_4^+ $-N/NH3-N assimilated by the isolate was 50.3 mg; so the ${\rm {NH}}_4^+ $-N/NH3-N removed by heterotrophic nitrification, in theory, is approximately 49.5 mg, which would result in a calculated heterotrophic nitrification rate of 0.69 mg/(L·h). There was a 40-fold difference between the two calculation methods. The actual nitrification capacity should be higher than the measured value because the strain also had denitrification capacity (Fig. 4). In addition, acetylene inhibits the oxidation of ${\rm {NH}}_4^+ $-N/NH3-N to ${\rm {NO}}_2^- $-N (Hynes and Knowles, 1978). The calculated heterotrophic nitrification rate of 0.69 mg/(L·h) is expected to be close to the actual value.
Figure
3.
The accumulation of ${\rm{NO}}_3^-$-N and ${\rm{NO}}_2^-$-N of strain H. venusta MA-ZP17-13 under heterotrophic nitrification condition at low temperature.
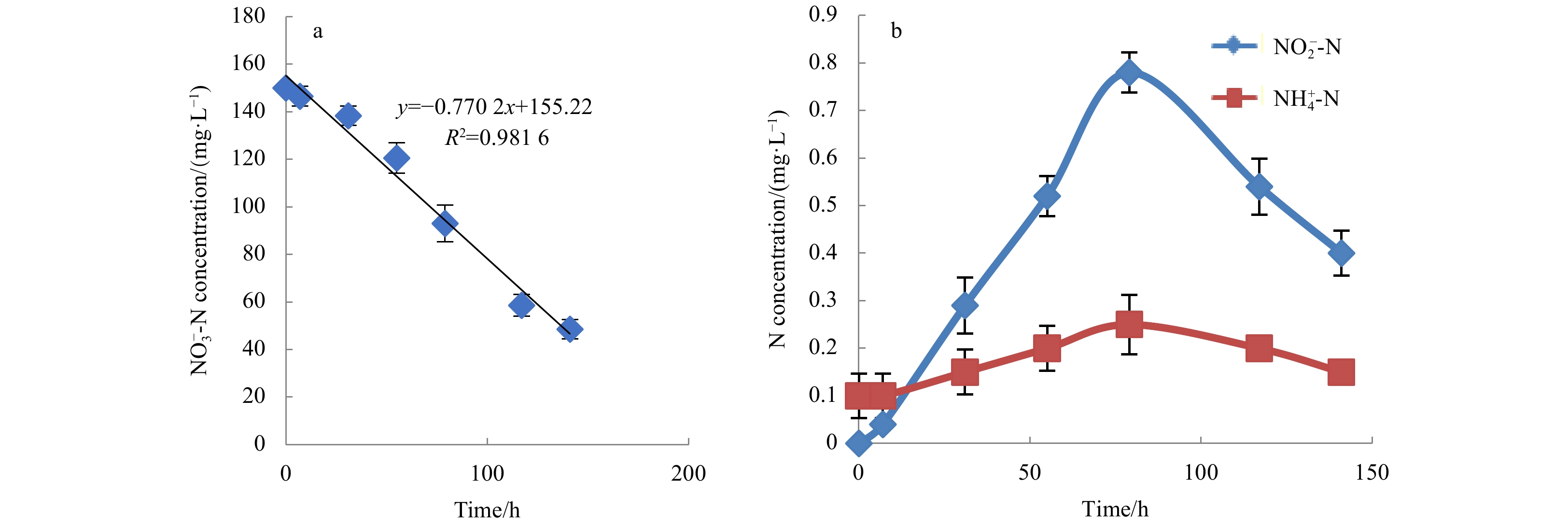
Figure
4.
The dynamic changes of ${\rm{NO}}_3^- $-N, ${\rm{NO}}_2^- $-N and ${\rm{NH}}_4^+$-N during denitrification process. a. Removal rate of ${\rm{NO}}_3^- $-N by strain MA-ZP17-13; b. the dynamic changes of${\rm{NO}}_2^- $-N and NH4+-N concentrations during denitrification.
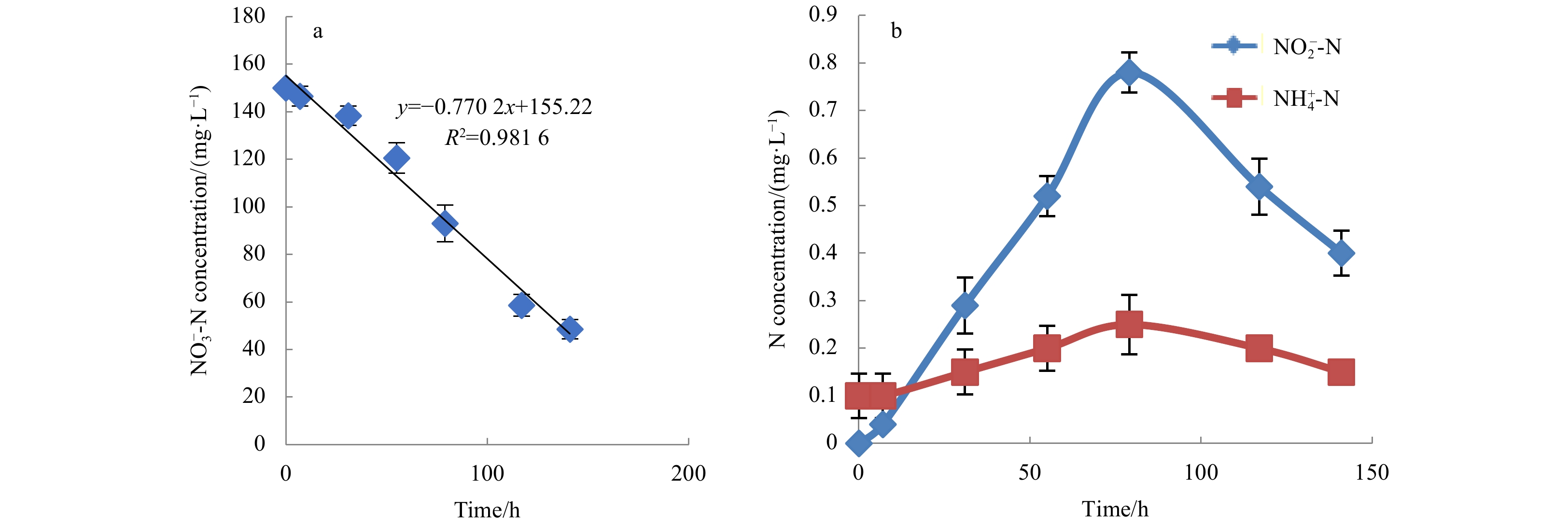
To determine its ${\rm {NO}}_3^- $-N removal capacity at 10°C, an initial concentration of 150 mg/L ${\rm {NO}}_3^- $-N was used as sole N source in DM medium. Incubation under aerobic conditions resulted in a significant decrease in the ${\rm {NO}}_3^- $-N concentration (Fig. 4a). The rate of decrease of ${\rm {NO}}_3^- $-N was determined by linear fitting, where the slope of linear fitting is the removal rate. The ${\rm {NO}}_3^- $-N removal rate of strain MA-ZP17-13 was 0.77 mg/(L·h). To clarify the denitrifying ability of strain MA-ZP17-13, changes of the ${\rm {NO}}_2^- $-N and ${\rm {NH}}_4^+ $-N/NH3-N concentrations were also detected; with the decrease in ${\rm {NO}}_3^- $-N concentration, the ${\rm {NO}}_2^- $-N concentration increased in the first 79 h, and then rapidly decreased to low concentration of about 0.3 mg/L (Fig. 4b). Unlike ${\rm {NO}}_2^- $-N, the ${\rm {NH}}_4^+ $-N/NH3-N concentrations was low and showed no significant change throughout the process. In addition, only N2O was observed as the end product of denitrification, no N2 was detected under aerobic conditions.
In conclusion, strain MA-ZP17-13 is capable of SND under low temperature. Compared with traditional denitrifying strains, SND has advantages such as high ${\rm {NH}}_4^+ $-N/NH3-N removal efficiency, fast microbial growth, and low process costs (Zhu et al., 2008; Wang et al., 2019). Therefore, the research on SND is of great significance to the sewage treatment industry.
3.3
Optimization of ammonia removal conditions
3.3.1
Development of regression model equations
Based on the single-factor experiments, a complete experimental design matrix was developed by BBD for further optimizations of the four parameters. The values of the response gained from the experiment were shown in Table 2. The ${\rm {NH}}_4^+ $-N/NH3-N removal percentage R1 ranged from 18.5% to 100.0%. For these responses, the empirical quadratic model was applied as recommendation for BBD to develop the correlation between the responses and independent variables as follows:
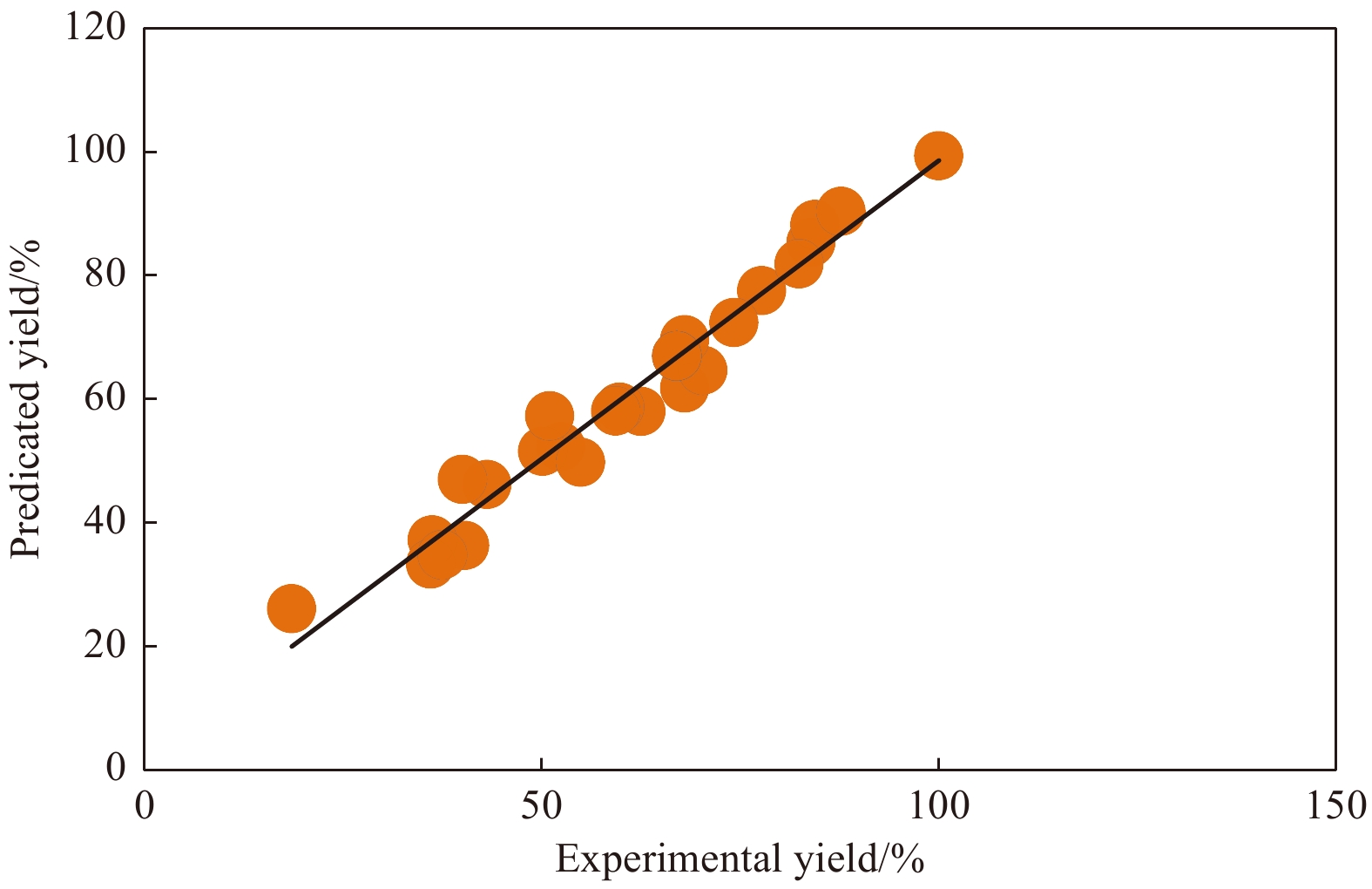
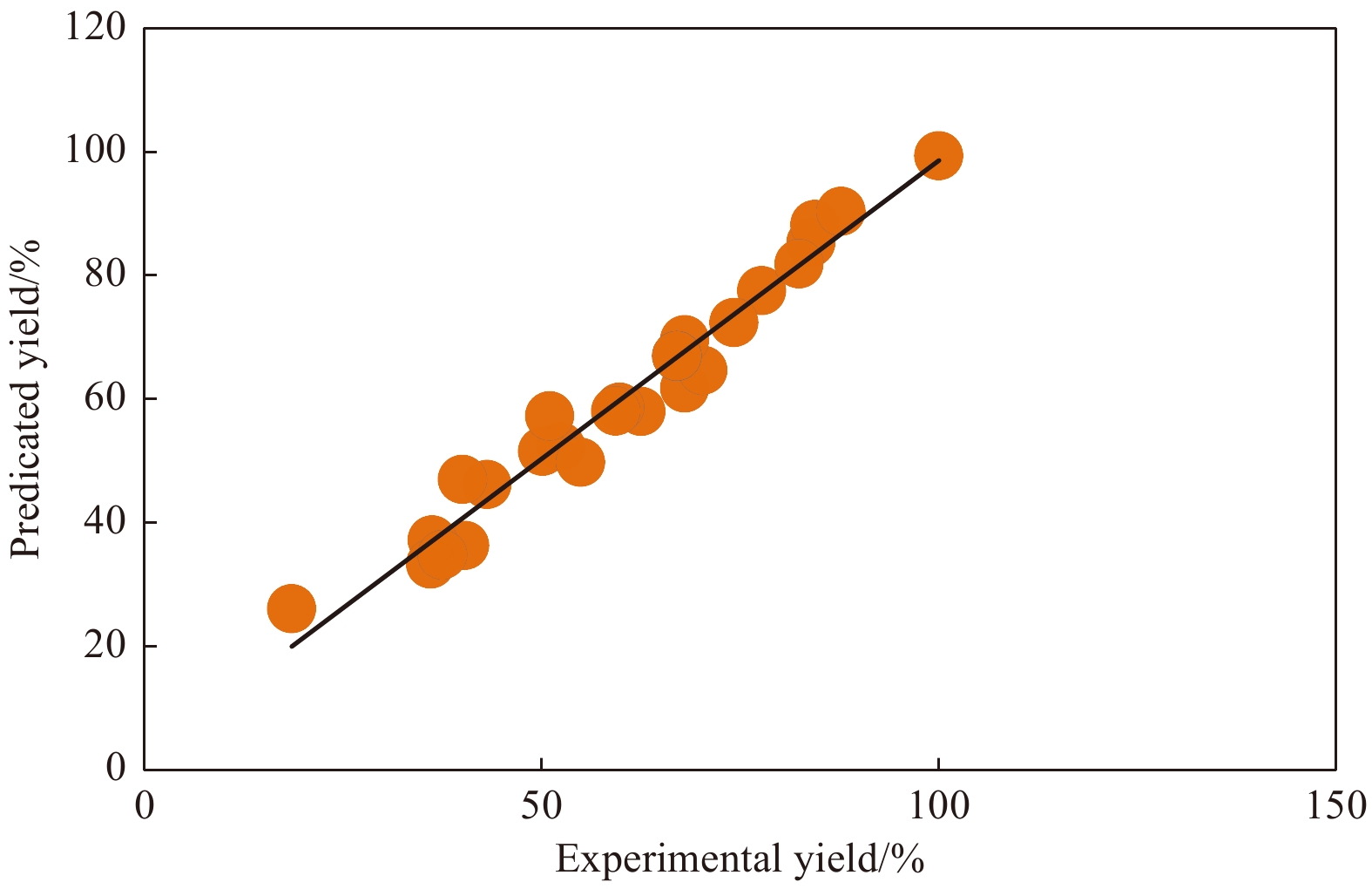
The accuracy of the established regression models was then assessed by the correlation coefficient R2, with values closer to 1, indicating a more precise response value estimated by the models. The R2 value for Eq. (1) was 0.96 and the predicted values versus the corresponding experimental values for Y1 were shown in Fig. 5. As expected, the actual values were consistent with the predicted values, implying that the models successfully predicted the relationships between culture parameters and ${\rm {NH}}_4^+ $-N/NH3-N removal.
Figure
5.
Predicted vs. experimental values of ammonia nitrogen removal.
In addition to the correlation coefficients, F-values and P-values were employed to determine the significance of the models and each model term, with a larger F-value indicating increased significance of the corresponding coefficient (Zhou et al., 2013). The ANOVA of the predicted quadratic model for ${\rm {NH}}_4^+ $-N/NH3-N removal (Table 4) showed that the model was statistically significant (F-value=27.57), indicating that most of the variations in the response could be explained with the regression equation. P<0.05 implied that the model terms were significant (Savasari et al., 2015). In this case, A (C/N concentration ratio), B (pH), C (NaCl), D (temperature), AB, AD, CD, the quadratic terms of salinity C2 and the quadratic terms of temperature D2 were significant.
Table
4.
ANOVA of the response surface quadratic model for ammonia nitrogen removal
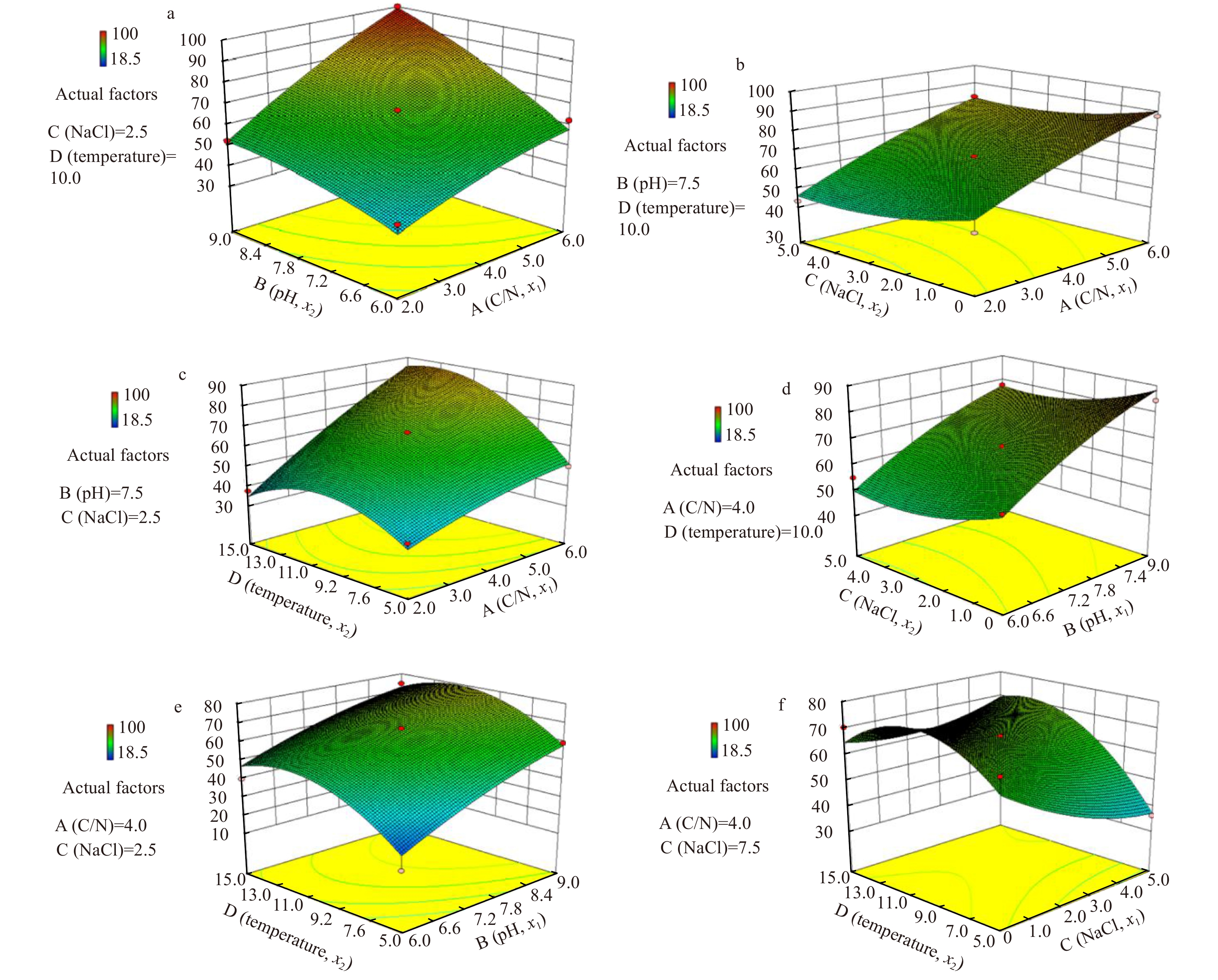
The ${\rm {NH}}_4^+ $-N/NH3-N removal capacities of strain MA-ZP17-13 for different combinations of the independent preparation parameters were visualized through 3D response surface plots (Fig. 6). The interactive effect between A (C/N concentration ratio) and B (pH) was observed (Fig. 6a), and confirmed by the corresponding F-value (6.65) and P (0.021 9; Table 4). The upward trend in ${\rm {NH}}_4^+ $-N/NH3-N removal was attributed to the increasing degree of A (C/N concentration ratio) and B (pH), with higher values of A and B resulting in higher ${\rm {NH}}_4^+ $-N/NH3-N removal rate. The interactive effect of A (C/N concentration ratio) and C (NaCl) on the ${\rm {NH}}_4^+ $-N/NH3-N removal rate was shown in Fig. 6b. The curved contour lines demonstrated that the increase in ${\rm {NH}}_4^+ $-N/NH3-N removal was attributed to the decrease in C (NaCl) and the increase in A (C/N concentration ratio). Similarly, a relatively strong interaction was identified between A (C/N concentration ratio) and D (temperature) on ${\rm {NH}}_4^+ $-N/NH3-N removal rate (Fig. 6c). During the initial stage, as these two factors increased, there is a simultaneous increase in the ${\rm {NH}}_4^+ $-N/NH3-N removal rate. Thereafter, a continued increase in D (temperature) lead to a reduction in the ${\rm {NH}}_4^+ $-N/NH3-N removal capacity, which indicates that strain MA-ZP17-13 is psychrophilic. There is a significant interactive effect between B (pH) and C (NaCl) on ${\rm {NH}}_4^+ $-N/NH3-N removal (Fig. 6d). The upward trend in ${\rm {NH}}_4^+ $-N/NH3-N removal was attributed to the slow decrease in C (NaCl) and the increase in B (pH). Salinity alone had little effect on ${\rm {NH}}_4^+ $/NH3 removal (confirmed by the corresponding F-value (11.92) and P (0.003 9) in (Table 4). There was an interactive effect between pH (B) and temperature (D) on ${\rm {NH}}_4^+ $-N/NH3-N removal (Fig. 6e). During the initial stage, as these two factors (NaCl and temperature) increased, ${\rm {NH}}_4^+ $-N/NH3-N removal increased. Thereafter, a continued increase in D (temperature) led to a slow downward trend in the ${\rm {NH}}_4^+ $-N/NH3-N removal capacity. Finally, there was an interactive effect between C (NaCl) and D (temperature) on ${\rm {NH}}_4^+ $-N/NH3-N removal (Fig. 6f). With the increase of these two factors, ${\rm {NH}}_4^+ $-N/NH3-N removal slowly increased, because the ${\rm {NH}}_4^+ $-N/NH3-N removal capacity was negatively correlated with concentration of NaCl (C). The higher the salinity, the lower the ${\rm {NH}}_4^+ $-N/NH3-N removal capacity. When C (NaCl) was 0, the increase in D (temperature) in the first stage resulted in increased ${\rm {NH}}_4^+ $-N/NH3-N removal capacity until the optimal level was reached; ${\rm {NH}}_4^+ $-N/NH3-N removal capacity then decreased with the increase in D (temperature) above the optimal value.
Figure
6.
The interactions and response surfaces of the culture parameters for the ammonia nitrogen removal.
The ${\rm {NH}}_4^+ $-N/NH3-N removal capacity was significantly influenced by four culture parameters of C/N ratio (A), pH (B), NaCl (C) and temperature (D). Although the influences of these four parameters were different, it was possible to balance the responses by searching for the optimum point. The optimal R1 was obtained using the following conditions: C/N ratio at 5.95, pH 8.93, NaCl at 2.33% and temperature at 11.2°C, which could result in 100% ${\rm {NH}}_4^+ $-N/NH3-N removal according to the regression models. Verification of the predictive results were carried out by three independent experiments under the predicted optimal conditions. The average experimental ${\rm {NH}}_4^+ $-N/NH3-N removal capacity (98.7%±0.9%) was similar to the values predicted from the models, with a small errors of 1.33%. Through process optimization, the ${\rm {NH}}_4^+ $-N/NH3-N removal rate was greatly increased from 88.0% to 98.7%, with the average ${\rm {NH}}_4^+ $-N/NH3-N removal rate of 1.37 mg/(L·h).
The ${\rm {NH}}_4^+ $-N/NH3-N removal ability of strain MA-ZP17-13 at low temperature was comparable to other previously reported bacteria, which were isolated from sediment, groundwater and soil, and the ammonia nitrogen removal rate were in the range of 0.092−3.03 mg/(L·h) (Table 5) (Huang et al., 2013; Yao et al., 2013; Qu et al., 2015). Compared with the above strains, strain MA-ZP17-13 had higher salinity tolerance, because it came from aquaculture seawater.
Table
5.
Comparison with other studies of ammonia nitrogen removal at low temperature
3.4
Genes and pathways involved nitrogen metabolisms based on the complete genomic sequencing
The complete genome sequence of strain MA-ZP17-13 was determined in this study. It consisted of one chromosome with a total length of 4 446 698 bp with a G+C content of 52.79% (according to amount of substance) (Fig. 7). Gene prediction identified 4 330 genes, of which 4 251 were CDSs, and 79 were RNAs (Table 6). The average length of the CDSs was 936 bp, giving a coding density of 96.0%. To further understand the adaptive capacity of strain MA-ZP17-13 to the aquaculture environment, metabolic features related to functional categories were analyzed. Twenty-five genes were found relating to N metabolism, which included nitrification and denitrification (Supplementary Table S1).
Figure
7.
Circular representation of Halomonas venusta MA-ZP17-13 genome. From the outside to the center: label of genome size (Mbp), CDSs on forward strand (colored by COG categories), CDSs on reverse strand (colored by COG categories), G+C content and GC skew. CDSs are depicted in different colors according to COG categories.
Genes encoding ammonia monooxygenase, nitric oxide dioxygenase, nitric oxide synthase, nitrite reductases, nitrate reductases and dissimilatory nitrite reductases were present in the genome of this strain, indicating that it was capable for both nitrification and denitrification. Consistent with its ${\rm {NH}}_4^+ $-N/NH3-N removal activity, the genome of H. venusta MA-ZP17-13 contained an ammonia monooxygenase gene (MA-ZP17-13_3189), which is 1 041 bp long and encodes 346 amino acids. Interestingly, the genome contained no hydroxylamine oxidase gene, which raises questions about the nitrification process of strain MA-ZP17-13. Nitric oxide dioxygenases (NODs) were present in the genome. NODs catalyze the reaction of O2 and NO to yield ${\rm {NO}}_3^- $ (Gardner et al., 1998; Lundberg et al., 2004; Schopfer et al., 2009). Therefore, this study inferred that there might be a new nitrification pathway in strain MA-ZP17-13, and the specific pathway was speculated as follows: ${\rm {NH}}_4^+ $→NH2OH→NO→${\rm {NO}}_3^- $.
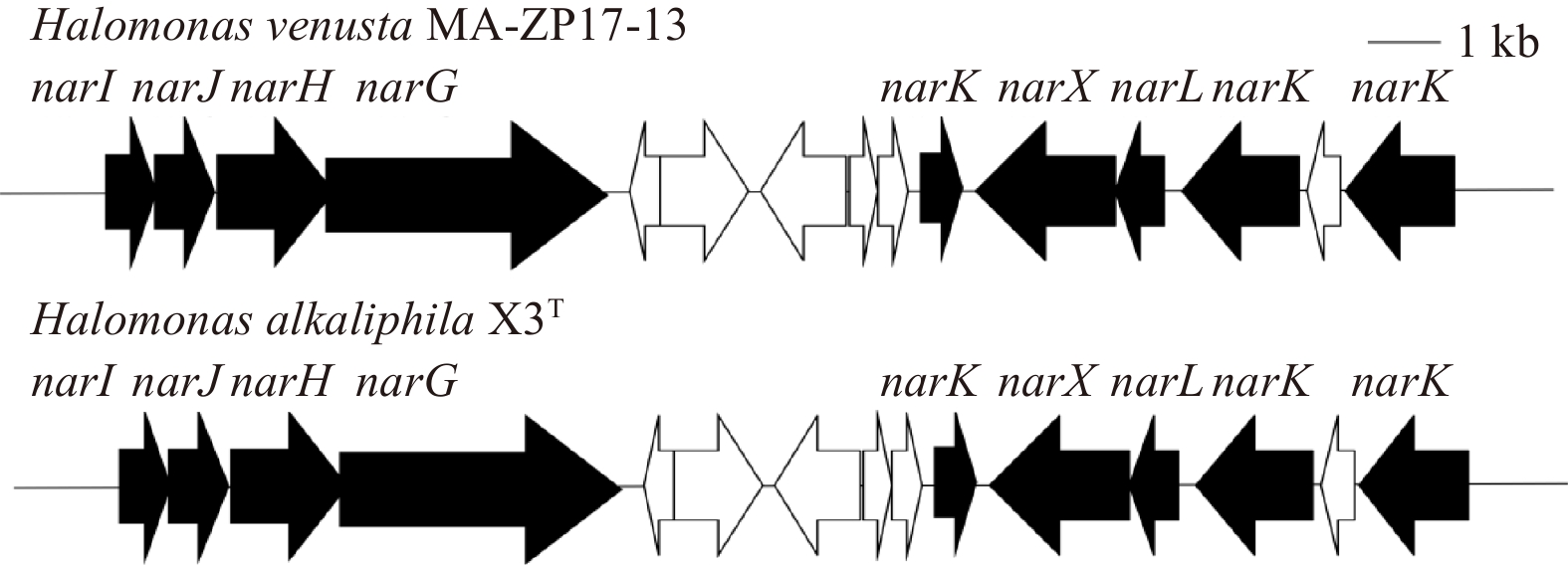
Consistent with the abilities of strain MA-ZP17-13, H. alkaliphila X3T was reported to be capable for heterotrophic nitrification-aerobic denitrification (Zhang et al., 2016) and the genome was obtained from NCBI database. These strains shared 99.68% similarity in 16S rRNA gene sequence, but the ANI between the two strains was 93.01%, indicating that the two strains belonged to two different species in genus Halomonas. This study found that H. alkaliphila X3T possessed similar nitrification-related genes and was also lacking hydroxylamine oxidase. The ammonia monooxygenase gene in strain MA-ZP17-13 is also 1 041 bp long and encodes 346 amino acids and is homologous to the corresponding gene in strain X3T with at least 99% amino acid identity. The high sequence similarity of the ammonia monooxygenase gene between strains MA-ZP17-13 and H. alkaliphila X3T indicates similar function and regulation of ${\rm {NH}}_4^+ $-N/NH3-N oxidation.
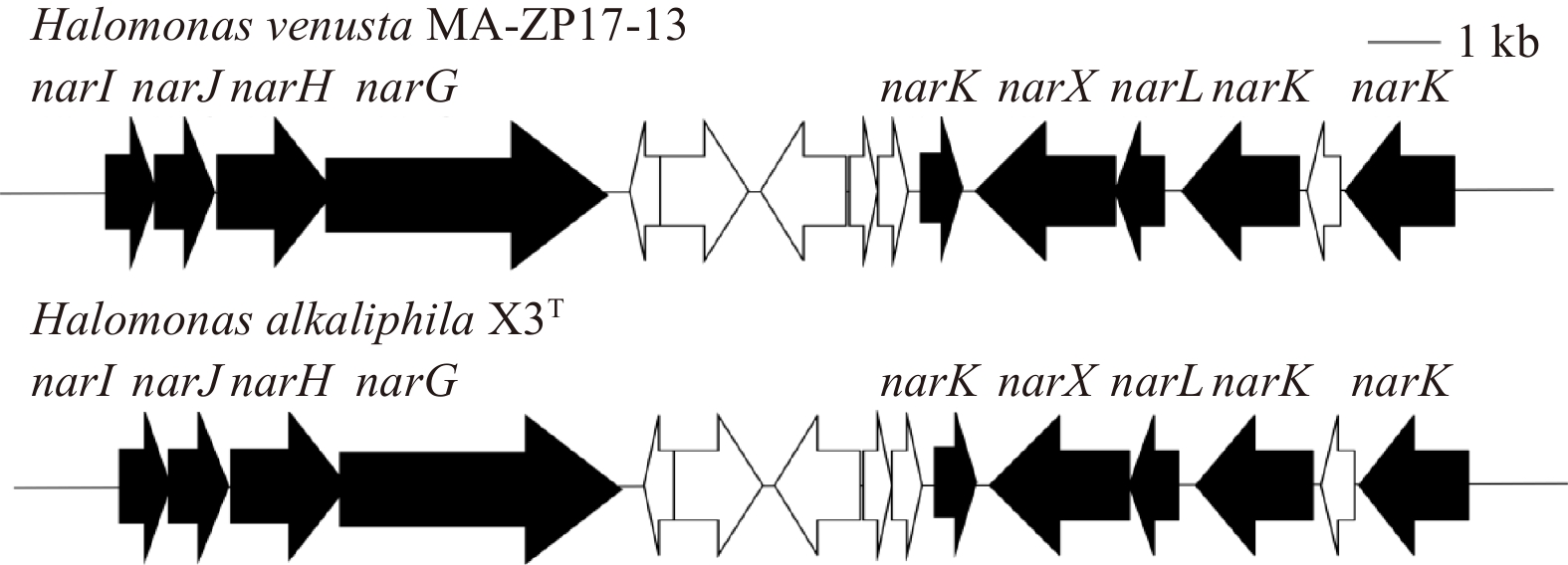
In addition, this study found that the same gene cluster of nitrate reductase was present in strains MA-ZP17-13 and H. alkaliphila X3T. The gene cluster contains the nitrate reductase gamma subunit (narI), nitrate reductase molybdenum cofactor assembly chaperone (narJ), nitrate reductase subunit beta (narH), nitrate reductase subunit alpha (narG), nitrate/nitrite sensor protein (narX), DNA-binding response regulator (narL), and MFS transporter (narK) genes (Fig. 8). Genes of nitrite reductases, nitrate reductases, nitric oxide dioxygenase, nitric oxide synthase, dissimilatory nitrite reductases were present in both strain MA-ZP17-13 and X3T. Validation of the function of these encoding genes requires further study to confirm their abilities and determine the mechanism of nitrification and denitrification, which is currently unknown. Increased understanding of the mechanisms of ${\rm {NH}}_4^+ $-N/NH3-N removal in this strain could aid in developing strategies for bioremediation of ${\rm {NH}}_4^+ $-N/NH3-N pollution.
The isolated Halomonas venusta MA-ZP17-13 is capable for ${\rm {NH}}_4^+ $-N/NH3-N removal via assimilation at 0.7 mg/(L·h), and also performs simultaneous heterotrophic nitrification and denitrification under aerobic conditions. Quantification results showed that about half of ${\rm {NH}}_4^+ $-N/NH3-N was removed by assimilation, and half was removed by denitrification firstly via nitrification. The optimal ammonia removal conditions were determined as C/N concentration ratio 5.95, pH 8.93, 2.33% NaCl and 11.2°C, which lead to 98.7% ${\rm {NH}}_4^+ $-N/NH3-N removed in 72 h at the initial concentration of 100 mg/L and the removal rate averaged at 1.37 mg/(L·h) (according to N). The ammonia removal potential is also supported by the presence of genes encoding ammonia monooxygenase, respiratory nitrate reductase, nitrite reductase, nitric oxide synthase and nitric oxide dioxygenase in the genome though the gene hydroxylamine oxidase was found in the complete genome sequence. This supposed a potential novel nitrification pathway of ${\rm {NH}}_4^+ $→NH2OH→NO→${\rm {NO}}_3^- $ and needs further investigation. Together, these results indicate that this bacterium has potential for ${\rm {NH}}_4^+ $-N/NH3-N removal in salty water at low temperature.
APHA. 2005. Standard Methods for the Examination of Water and Wastewater. 21th ed. Washington, DC, USA: American Public Health Association (APHA)
[2]
Armstrong D A, Chippendale D, Knight A W, et al. 1978. Interaction of ionized and un-ionized ammonia on short-term survival and growth of prawn larvae, Macrobrachium rosenbergh. The Biological Bulletin, 154(1): 15–31. doi: 10.2307/1540771
[3]
Atanassov C L, Muller C D, Sarhan S, et al. 1994. Effect of ammonia on endocytosis, cytokine production and lysosomal enzyme activity of a microglial cell line. Research in Immunology, 145(4): 277–288. doi: 10.1016/S0923-2494(94)80016-2
[4]
Auch A F, Klenk H P, Göker M. 2010a. Standard operating procedure for calculating genome-to-genome distances based on high-scoring segment pairs. Standards in Genomic Sciences, 2(1): 142–148. doi: 10.4056/sigs.541628
[5]
Auch A F, Von Jan M, Klenk H P, et al. 2010b. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Standards in Genomic Sciences, 2(1): 117–134. doi: 10.4056/sigs.531120
[6]
Carrera J, Vicent T, Lafuente F J. 2003. Influence of temperature on denitrification of an industrial high-strength nitrogen wastewater in a two-sludge system. Water SA, 29(1): 11–16
[7]
Chen Qian, Ni Jinren. 2012. Ammonium removal by Agrobacterium sp. LAD9 capable of heterotrophic nitrification–aerobic denitrification. Journal of Bioscience and Bioengineering, 113(5): 619–623. doi: 10.1016/j.jbiosc.2011.12.012
[8]
Chin C S, Alexander D H, Marks P, et al. 2013. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nature Methods, 10(6): 563–569. doi: 10.1038/nmeth.2474
[9]
Chiu Y C, Lee L L, Chang Chengnan, et al. 2007. Control of carbon and ammonium ratio for simultaneous nitrification and denitrification in a sequencing batch bioreactor. International Biodeterioration & Biodegradation, 59(1): 1–7
[10]
Gardner P R, Gardner A M, Martin L A, et al. 1998. Nitric oxide dioxygenase: an enzymic function for flavohemoglobin. Proceedings of the National Academy of Sciences of the United States of America, 95(18): 10378–10383. doi: 10.1073/pnas.95.18.10378
[11]
Goris J, Konstantinidis K T, Klappenbach J A, et al. 2007. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. International Journal of Systematic and Evolutionary Microbiology, 57(1): 81–91. doi: 10.1099/ijs.0.64483-0
[12]
He Tengxia, Li Zhenlun, Sun Quan, et al. 2016. Heterotrophic nitrification and aerobic denitrification by Pseudomonas tolaasii Y-11 without nitrite accumulation during nitrogen conversion. Bioresource Technology, 200: 493–499. doi: 10.1016/j.biortech.2015.10.064
[13]
He Tengxia, Xie Deti, Li Zhenlun, et al. 2017. Ammonium stimulates nitrate reduction during simultaneous nitrification and denitrification process by Arthrobacter arilaitensis Y-10. Bioresource Technology, 239: 66–73. doi: 10.1016/j.biortech.2017.04.125
[14]
Hooper A B, Vannelli T, Bergmann D J, et al. 1997. Enzymology of the oxidation of ammonia to nitrite by bacteria. Antonie van Leeuwenhoek, 71(1): 59–67
[15]
Huang Xiaofei, Li Weiguang, Zhang Duoying, et al. 2013. Ammonium removal by a novel oligotrophic Acinetobacter sp. Y16 capable of heterotrophic nitrification-aerobic denitrification at low temperature. Bioresource Technology, 146: 44–50. doi: 10.1016/j.biortech.2013.07.046
[16]
Hynes R K, Knowles R. 1978. Inhibition by acetylene of ammonia oxidation in Nitrosomonas europaea. FEMS Microbiology Letters, 4(6): 319–321. doi: 10.1111/j.1574-6968.1978.tb02889.x
[17]
Kan Fu, Xia Qinbin, Li Zhong, et al. 2011. Research of ammonia adsorption with Maifan stone. Industrial Safety and Environmental Protection, 37(4): 3–5
[18]
Kim J K, Park K J, Cho K S, et al. 2005. Aerobic nitrification–denitrification by heterotrophic Bacillus strains. Bioresource Technology, 96(17): 1897–1906. doi: 10.1016/j.biortech.2005.01.040
[19]
Koren S, Schatz M C, Walenz B P, et al. 2012. Hybrid error correction and de novo assembly of single-molecule sequencing reads. Nature Biotechnology, 30(7): 693–700. doi: 10.1038/nbt.2280
[20]
Krzywinski M, Schein J, Birol I, et al. 2009. Circos: an information aesthetic for comparative genomics. Genome Research, 19(9): 1639–1645. doi: 10.1101/gr.092759.109
[21]
Li Guizhen, Lai Qiliang, Yan Peisheng, et al. 2019. Roseovarius amoyensis sp. nov. and Muricauda amoyensis sp. nov., isolated from the Xiamen coast. International Journal of Systematic and Evolutionary Microbiology, 69(10): 3100–3108. doi: 10.1099/ijsem.0.003595
[22]
Lin Y C, Chen J C. 2003. Acute toxicity of nitrite on Litopenaeus vannamei (Boone) juveniles at different salinity levels. Aquaculture, 224(1–4): 193–201. doi: 10.1016/S0044-8486(03)00220-5
[23]
Lu Lu, Jia Zhongjun. 2013. Urease gene-containing Archaea dominate autotrophic ammonia oxidation in two acid soils. Environmental Microbiology, 15(6): 1795–1809. doi: 10.1111/1462-2920.12071
[24]
Lundberg J O, Weitzberg E, Cole J A, et al. 2004. Nitrate, bacteria and human health. Nature Reviews Microbiology, 2(7): 593–602. doi: 10.1038/nrmicro929
[25]
McCarty G W. 1999. Modes of action of nitrification inhibitors. Biology and Fertility of Soils, 29(1): 1–9. doi: 10.1007/s003740050518
[26]
Meier-Kolthoff J P, Auch A F, Klenk H P, et al. 2013. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics, 14: 60. doi: 10.1186/1471-2105-14-60
[27]
Myers R H, Montgomery D C, Anderson-Cook C M. 2016. Response Surface Methodology: Process and Product Optimization Using Designed Experiments. New York: John Wiley & Sons
[28]
Qu Dan, Wang Cong, Wang Yanfang, et al. 2015. Heterotrophic nitrification and aerobic denitrification by a novel groundwater origin cold-adapted bacterium at low temperatures. RSC Advances, 5(7): 5149–5157. doi: 10.1039/C4RA13141J
[29]
Qu Jianhua, Meng Xianlin, You Hong, et al. 2017. Utilization of rice husks functionalized with xanthates as cost-effective biosorbents for optimal Cd(II) removal from aqueous solution via response surface methodology. Bioresource Technology, 241: 1036–1042. doi: 10.1016/j.biortech.2017.06.055
[30]
Ren Yongxiang, Yang Lei, Liang Xian. 2014. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB. Bioresource Technology, 171: 1–9. doi: 10.1016/j.biortech.2014.08.058
[31]
Richter M, Rosselló-Móra R. 2009. Shifting the genomic gold standard for the prokaryotic species definition. Proceedings of the National Academy of Sciences of the United States of America, 106(45): 19126–19131. doi: 10.1073/pnas.0906412106
[32]
Rodriguez-Caballero A, Hallin S, Påhlson C, et al. 2012. Ammonia oxidizing bacterial community composition and process performance in wastewater treatment plants under low temperature conditions. Water Science and Technology, 65(2): 197–204. doi: 10.2166/wst.2012.643
[33]
Ryden J C, Skinner J H, Nixon D J. 1987. Soil core incubation system for the field measurement of denitrification using acetylene-inhibition. Soil Biology and Biochemistry, 19(6): 753–757. doi: 10.1016/0038-0717(87)90059-9
[34]
Savasari M, Emadi M, Ali Bahmanyar M, et al. 2015. Optimization of Cd (II) removal from aqueous solution by ascorbic acid-stabilized zero valent iron nanoparticles using response surface methodology. Journal of Industrial and Engineering Chemistry, 21: 1403–1409. doi: 10.1016/j.jiec.2014.06.014
[35]
Schopfer M P, Mondal B, Lee D H, et al. 2009. Heme/O2/·NO Nitric oxide dioxygenase (NOD) reactivity: phenolic nitration via a putative heme-peroxynitrite intermediate. Journal of the American Chemical Society, 131(32): 11304–11305. doi: 10.1021/ja904832j
[36]
Schuler D J, Boardman G D, Kuhn D D, et al. 2010. Acute toxicity of ammonia and nitrite to pacific white shrimp, Litopenaeus vannamei, at low salinities. Journal of the World Aquaculture Society, 41(3): 438–446. doi: 10.1111/j.1749-7345.2010.00385.x
[37]
Shan H, Obbard J. 2001. Ammonia removal from prawn aquaculture water using immobilized nitrifying bacteria. Applied Microbiology and Biotechnology, 57(5−6): 791–798. doi: 10.1007/s00253-001-0835-1
[38]
Smart G R. 1978. Investigations of the toxic mechanisms of ammonia to fish–gas exchange in rainbow trout (Salmo gairdneri) exposed to acutely lethal concentrations. Journal of Fish Biology, 12(1): 93–104. doi: 10.1111/j.1095-8649.1978.tb04155.x
[39]
Tatusova T, Dicuccio M, Badretdin A, et al. 2016. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Research, 44(14): 6614–6624. doi: 10.1093/nar/gkw569
[40]
Thurston R V, Russo R C, Vinogradov G A. 1981. Ammonia toxicity to fishes. Effect of pH on the toxicity of the unionized ammonia species. Environmental Science & Technology, 15(7): 837–840
[41]
Wang Te, Jiang Zhengzhong, Dong Wenbo, et al. 2019. Growth and nitrogen removal characteristics of Halomonas sp. B01 under high salinity. Annals of Microbiology, 69(13): 1425–1433. doi: 10.1007/s13213-019-01526-y
[42]
Wayne L G, Brenner D J, Colwell R R, et al. 1987. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. International Journal of Systematic and Evolutionary Microbiology, 37(4): 463–464. doi: 10.1099/00207713-37-4-463
[43]
Wei Yunxia, Li Yanfeng, Ye Zhengfang. 2010. Enhancement of removal efficiency of ammonia nitrogen in sequencing batch reactor using natural zeolite. Environmental Earth Sciences, 60(7): 1407–1413. doi: 10.1007/s12665-009-0276-1
[44]
Xu Yi, He Tengxia, Li Zhenlun, et al. 2017. Nitrogen removal characteristics of Pseudomonas putida Y-9 capable of heterotrophic nitrification and aerobic denitrification at low temperature. BioMed Research International, 2017: 1429018
[45]
Yao Shuo, Ni Jinren, Ma Tao, et al. 2013. Heterotrophic nitrification and aerobic denitrification at low temperature by a newly isolated bacterium, Acinetobacter sp. HA2. Bioresource Technology, 139: 80–86. doi: 10.1016/j.biortech.2013.03.189
[46]
Yu Lei, Wang Yangqing, Liu Hongjie, et al. 2016. A novel heterotrophic nitrifying and aerobic denitrifying bacterium, Zobellella taiwanensis DN-7, can remove high-strength ammonium. Applied Microbiology and Biotechnology, 100(9): 4219–4229. doi: 10.1007/s00253-016-7290-5
[47]
Zerbino D R. 2010. Using the Velvet de novo assembler for short-read sequencing technologies. Current Protocols in Bioinformatics, 31(1): 5–11
[48]
Zhang Yan, Cheng Yu, Fei Yutao, et al. 2016. Response to different nitrogen forms of heterotrophic nitrifying-aerobic denitrifying bacteria X3. Advances in Marine Sciences, 3(4): 118–126. doi: 10.12677/AMS.2016.34016
[49]
Zhang Duoying, Li Weiguang, Huang Xiaofei, et al. 2013. Removal of ammonium in surface water at low temperature by a newly isolated Microbacterium sp. strain SFA13. Bioresource Technology, 137: 147–152. doi: 10.1016/j.biortech.2013.03.094
[50]
Zhang Jinbo, Sun Weijun, Zhong Wenhui, et al. 2014. The substrate is an important factor in controlling the significance of heterotrophic nitrification in acidic forest soils. Soil Biology and Biochemistry, 76: 143–148. doi: 10.1016/j.soilbio.2014.05.001
[51]
Zhang Jibin, Wu Pengxia, Hao Bo, et al. 2011. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001. Bioresource Technology, 102(21): 9866–9869. doi: 10.1016/j.biortech.2011.07.118
[52]
Zhao Bin, An Qiang, He Yiliang, et al. 2012. N2O and N2 production during heterotrophic nitrification by Alcaligenes faecalis strain NR. Bioresource Technology, 116: 379–385. doi: 10.1016/j.biortech.2012.03.113
[53]
Zhao Bin, He Yiliang, Huang Jue, et al. 2010b. Heterotrophic nitrogen removal by Providencia rettgeri strain YL. Journal of Industrial Microbiology & Biotechnology, 37(6): 609–616
[54]
Zhao Bin, He Yiliang, Hughes J, et al. 2010a. Heterotrophic nitrogen removal by a newly isolated Acinetobacter calcoaceticus HNR. Bioresource Technology, 101(14): 5194–5200. doi: 10.1016/j.biortech.2010.02.043
[55]
Zhou Xiang, Xin Zhijun, Lu Xihong, et al. 2013. High efficiency degradation crude oil by a novel mutant irradiated from Dietzia strain by 12C6+ heavy ion using response surface methodology. Bioresource Technology, 137: 386–393. doi: 10.1016/j.biortech.2013.03.097
[56]
Zhu Guibing, Peng Yongzhen, Li Baikun, et al. 2008. Biological removal of nitrogen from wastewater. In: Whitacre D M, ed. Reviews of Environmental Contamination and Toxicology. New York: Springer, 192: 159–195
Zhao Chen, Jian Li, Qianqian Zhai, et al. Nitrogen cycling process and application in different prawn culture modes. Reviews in Aquaculture, 2024, 16(4): 1580. doi:10.1111/raq.12912
2.
Ling Wang, You-Wei Cui. Mutualistic symbiosis of fungi and nitrogen-fixing bacteria in halophilic aerobic granular sludge treating nitrogen-deficient hypersaline organic wastewater. Bioresource Technology, 2024, 394: 130183. doi:10.1016/j.biortech.2023.130183
3.
Elizaveta P. Pulikova, Andrey V. Gorovtsov, Yakov Kuzyakov, et al. Heterotrophic nitrification in soils: approaches and mechanisms. Soil Biology and Biochemistry, 2024. doi:10.1016/j.soilbio.2024.109706
4.
Keke Lei, Zhaohua Wang, Shen Ma, et al. Construction of suspended bagasse bioflocs and evaluation of their effectiveness in shrimp (Penaeus vannamei) aquaculture systems. Aquaculture, 2024, 586: 740826. doi:10.1016/j.aquaculture.2024.740826
5.
Rohini Mattoo, Suman B M. Microbial roles in the terrestrial and aquatic nitrogen cycle—implications in climate change. FEMS Microbiology Letters, 2023, 370 doi:10.1093/femsle/fnad061
6.
Yumeng Xie, Xiangli Tian, Yu He, et al. Nitrogen removal capability and mechanism of a novel heterotrophic nitrification–aerobic denitrification bacterium Halomonas sp. DN3. Bioresource Technology, 2023, 387: 129569. doi:10.1016/j.biortech.2023.129569
7.
Guizhen Li, Mengjiao Wei, Guangshan Wei, et al. Efficient heterotrophic nitrification by a novel bacterium Sneathiella aquimaris 216LB-ZA1-12T isolated from aquaculture seawater. Ecotoxicology and Environmental Safety, 2023, 266: 115588. doi:10.1016/j.ecoenv.2023.115588
8.
Yuchen Yuan, Jiadong Liu, Bo Gao, et al. The Effect of Activated Sludge Treatment and Catalytic Ozonation on High Concentration of Ammonia Nitrogen Removal from Landfill Leachate. SSRN Electronic Journal, 2022. doi:10.2139/ssrn.4151696
9.
Yuchen Yuan, Jiadong Liu, Bo Gao, et al. The effect of activated sludge treatment and catalytic ozonation on high concentration of ammonia nitrogen removal from landfill leachate. Bioresource Technology, 2022, 361: 127668. doi:10.1016/j.biortech.2022.127668
10.
Yiming Yan, Hongwei Lu, Jin Zhang, et al. Simultaneous heterotrophic nitrification and aerobic denitrification (SND) for nitrogen removal: A review and future perspectives. Environmental Advances, 2022, 9: 100254. doi:10.1016/j.envadv.2022.100254
Figure 1. Neighbour-joining tree showing the phylogenetic positions of strain MA-ZP17-13 and other members of the genus Halomonas, based on 16S rRNA gene sequences. The bootstrap values on the branching nodes were calculated on 1 000 replications. The scale bar indicated 0.005 substitutions per nucleotide position.
Figure 2. Ammonium-nitrogen removal rate of strain Halomonas venusta MA-ZP17-13.
Figure 3. The accumulation of ${\rm{NO}}_3^-$-N and ${\rm{NO}}_2^-$-N of strain H. venusta MA-ZP17-13 under heterotrophic nitrification condition at low temperature.
Figure 4. The dynamic changes of ${\rm{NO}}_3^- $-N, ${\rm{NO}}_2^- $-N and ${\rm{NH}}_4^+$-N during denitrification process. a. Removal rate of ${\rm{NO}}_3^- $-N by strain MA-ZP17-13; b. the dynamic changes of${\rm{NO}}_2^- $-N and NH4+-N concentrations during denitrification.
Figure 5. Predicted vs. experimental values of ammonia nitrogen removal.
Figure 6. The interactions and response surfaces of the culture parameters for the ammonia nitrogen removal.
Figure 7. Circular representation of Halomonas venusta MA-ZP17-13 genome. From the outside to the center: label of genome size (Mbp), CDSs on forward strand (colored by COG categories), CDSs on reverse strand (colored by COG categories), G+C content and GC skew. CDSs are depicted in different colors according to COG categories.











































 DownLoad:
DownLoad:
 DownLoad:
DownLoad:


























































































 DownLoad:
DownLoad: