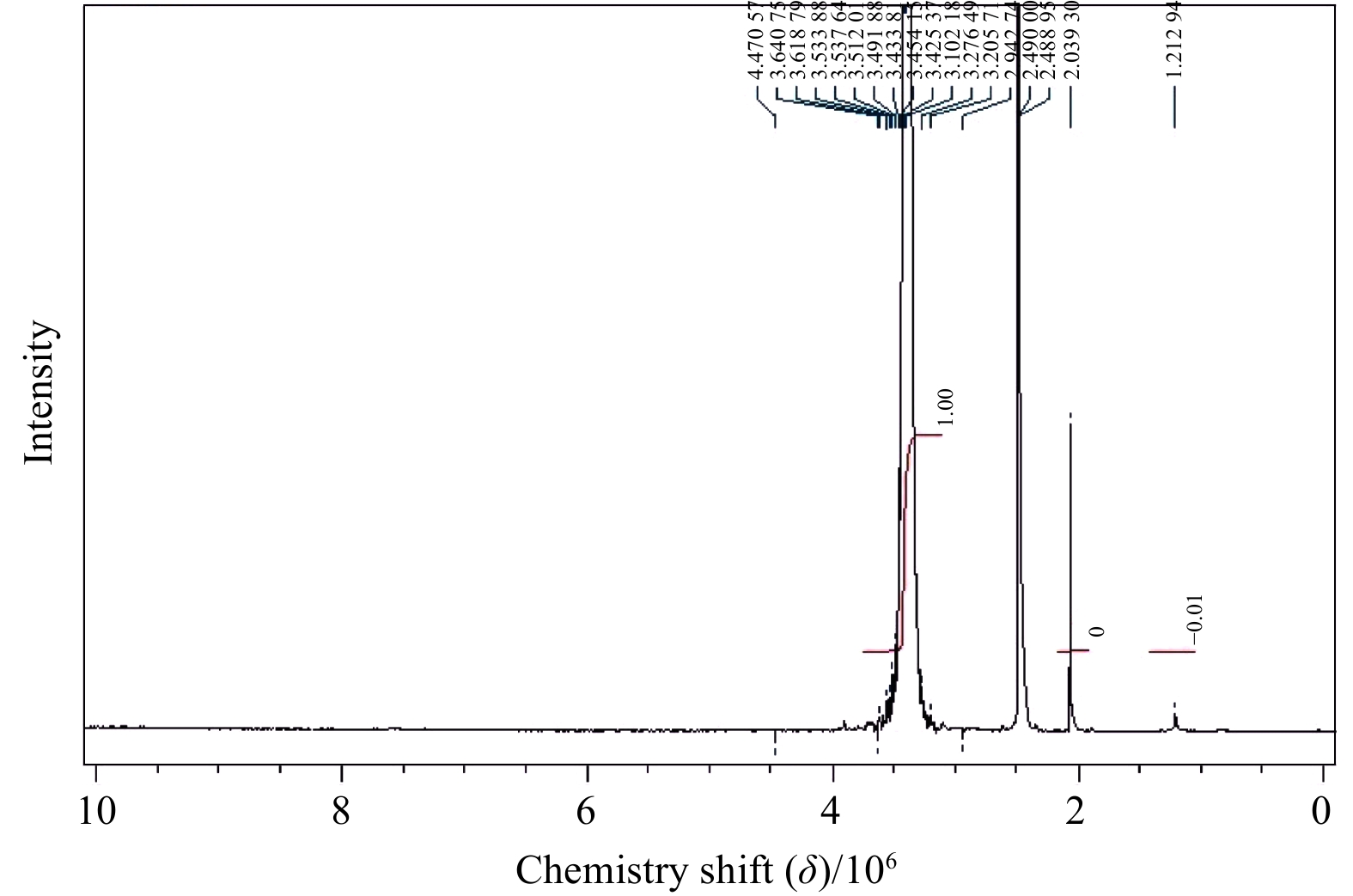
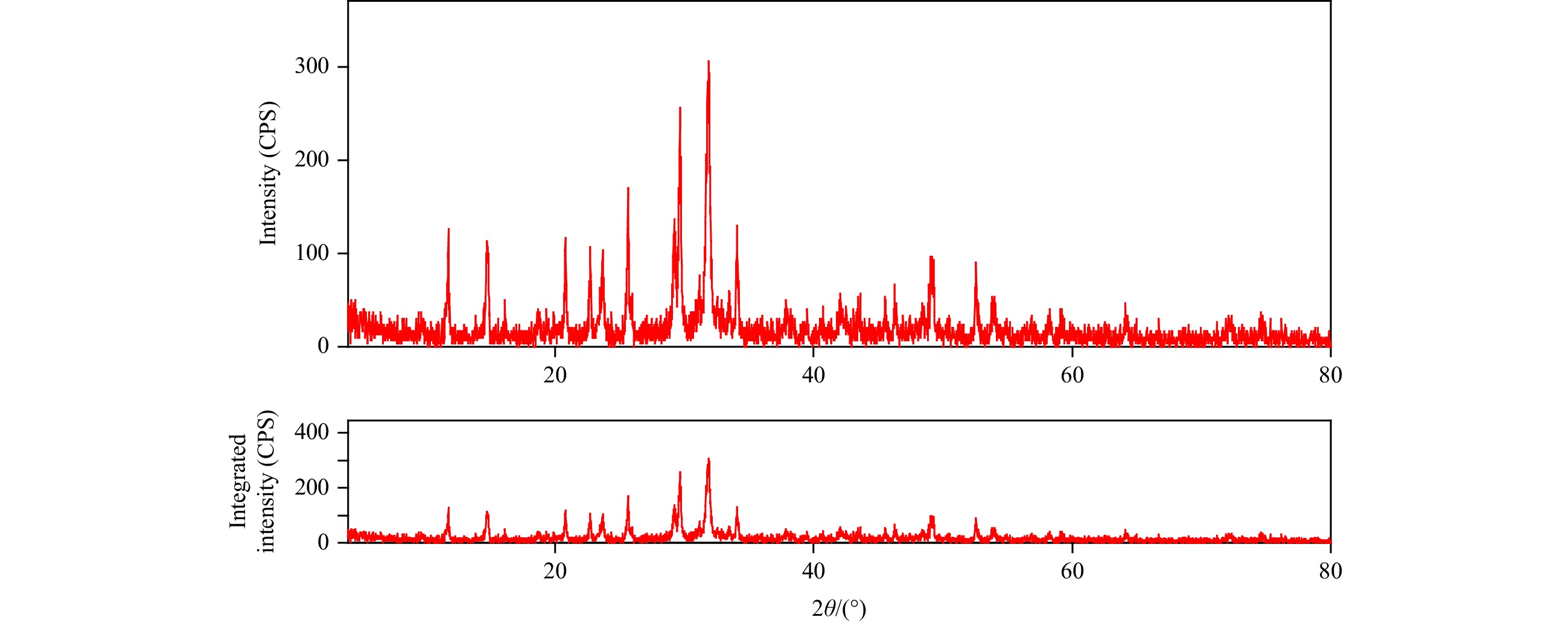
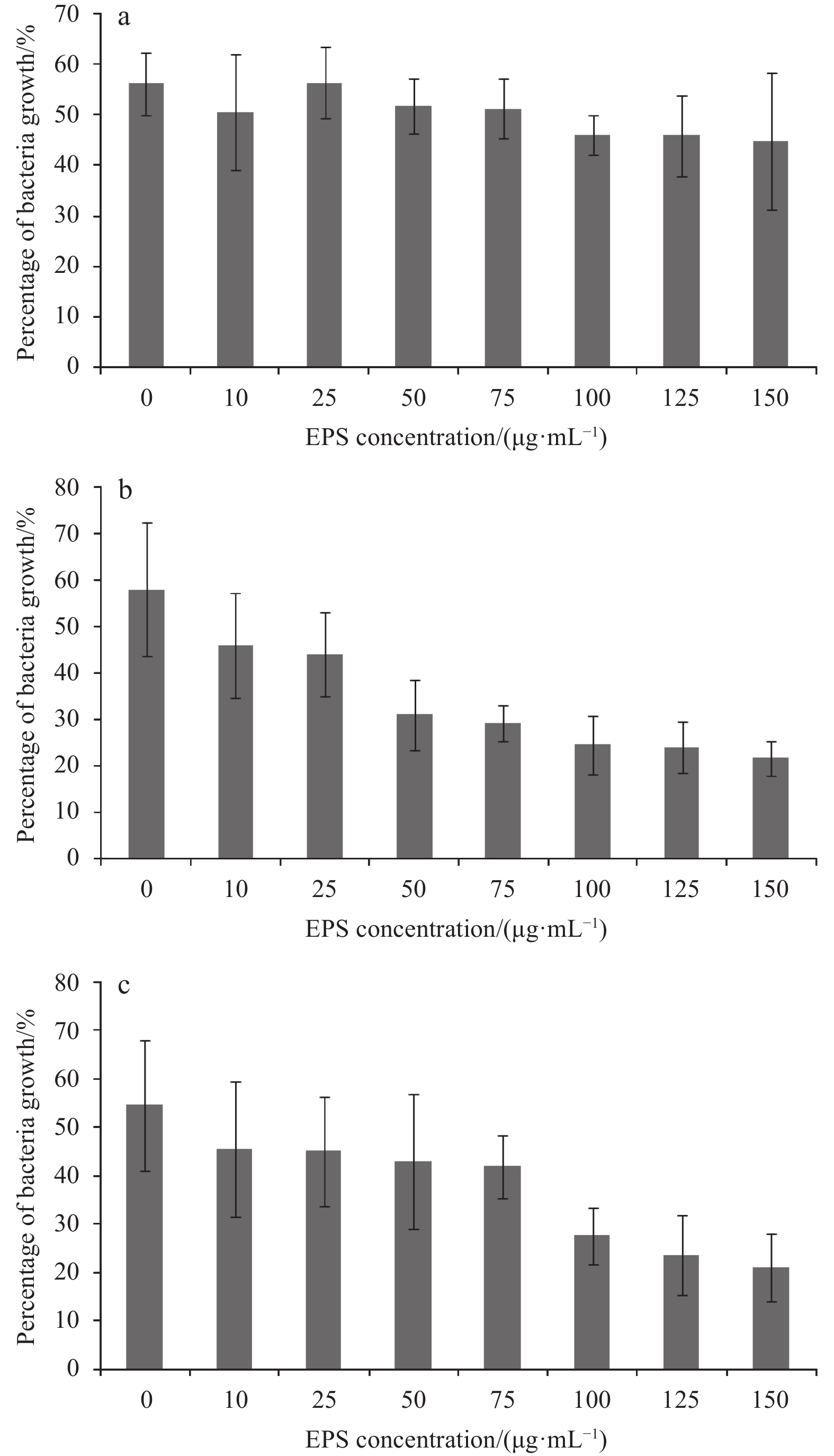
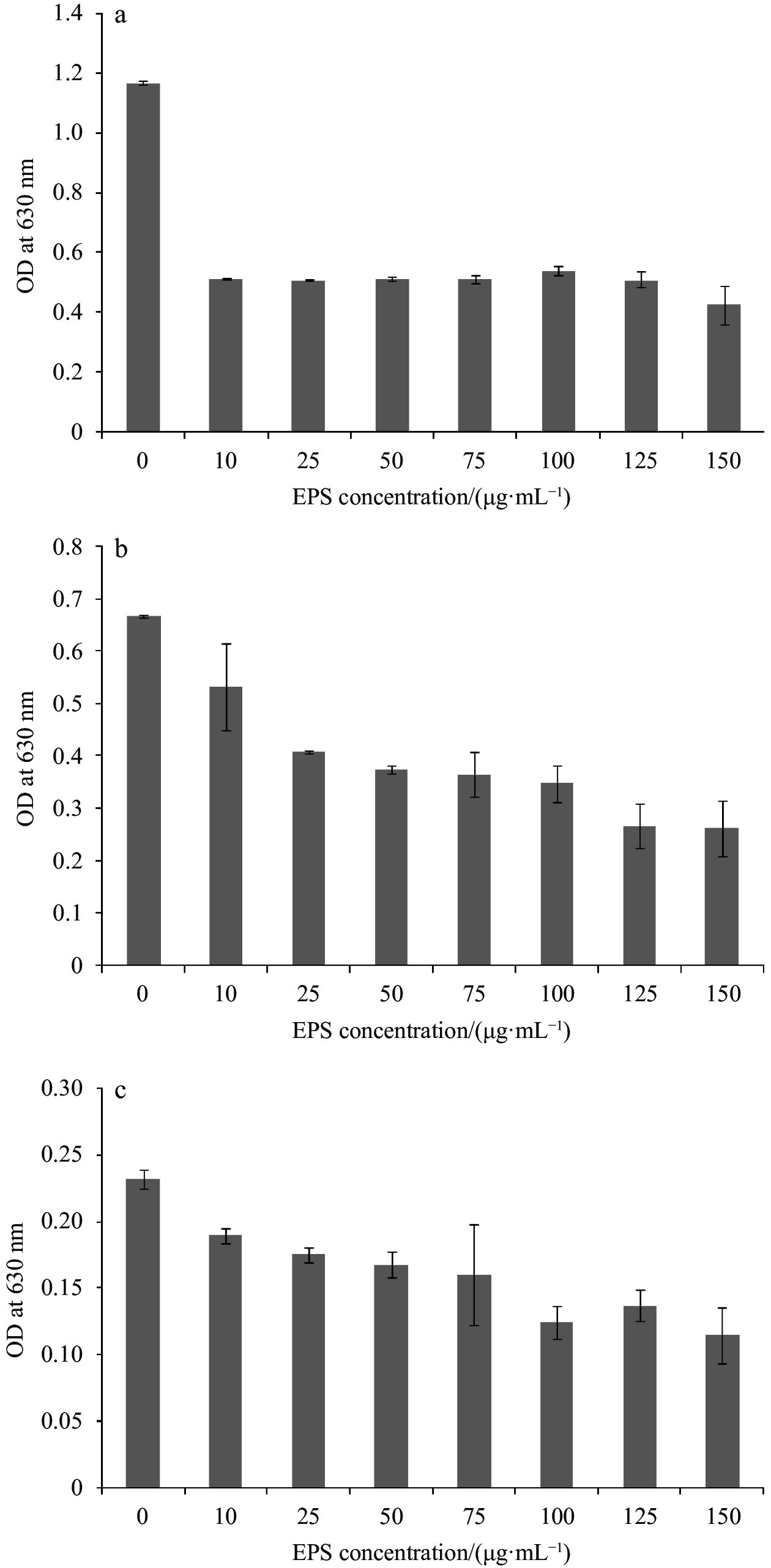
| Citation: | Mohammad Abdulaziz Ba-akdah, Sathianeson Satheesh. Characterization and antifouling activity analysis of extracellular polymeric substances produced by an epibiotic bacterial strain Kocuria flava associated with the green macroalga Ulva lactuca[J]. Acta Oceanologica Sinica, 2021, 40(4): 107-115. doi: 10.1007/s13131-020-1694-x

|

| [1] |
Almeida J R, Correia-da-Silva M, Sousa E, et al. 2017. Antifouling potential of nature-inspired sulfated compounds. Scientific Reports, 7: 42424. doi: 10.1038/srep42424
|
| [2] |
Angelina, Vijayendra S V N. 2015. Microbial biopolymers: the exopolysaccharides. In: Kalia V C, ed. Microbial Factories. New Delhi: Springer, 113–125
|
| [3] |
Badireddy A R, Korpol B R, Chellam S, et al. 2008. Spectroscopic characterization of extracellular polymeric substances from Escherichia coli and Serratia marcescens: suppression using sub-inhibitory concentrations of bismuth thiols. Biomacromolecules, 9(11): 3079–3089. doi: 10.1021/bm800600p
|
| [4] |
Balqadi A A, Salama A J, Satheesh S. 2018. Microfouling development on artificial substrates deployed in the central Red Sea. Oceanologia, 60(2): 219–231. doi: 10.1016/j.oceano.2017.10.006
|
| [5] |
Bérdy J. 2005. Bioactive microbial metabolites. The Journal of Antibiotics, 58(1): 1–26. doi: 10.1038/ja.2005.1
|
| [6] |
Bibi F, Naseer M I, Hassan A M, et al. 2018. Diversity and antagonistic potential of bacteria isolated from marine grass Halodule uninervis. 3 Biotech, 8(1): 48. doi: 10.1007/s13205-017-1066-1
|
| [7] |
Brian-Jaisson F, Molmeret M, Fahs A, et al. 2016. Characterization and anti-biofilm activity of extracellular polymeric substances produced by the marine biofilm-forming bacterium Pseudoalteromonas ulvae strain TC14. Biofouling, 32(5): 547–560. doi: 10.1080/08927014.2016.1164845
|
| [8] |
Camacho-Chab J C, Lango-Reynoso F, Castañeda-Chávez M D R, et al. 2016. Implications of extracellular polymeric substance matrices of microbial habitats associated with coastal aquaculture systems. Water, 8(9): 369. doi: 10.3390/w8090369
|
| [9] |
Caruso C, Rizzo C, Mangano S, et al. 2018. Production and biotechnological potential of extracellular polymeric substances from sponge-associated Antarctic bacteria. Applied and Environmental Microbiology, 84(4): e01624–17. doi: 10.1128/AEM.01624-17
|
| [10] |
Casillo A, Lanzetta R, Parrilli M, et al. 2018. Exopolysaccharides from marine and marine extremophilic bacteria: structures, properties, ecological roles and applications. Marine Drugs, 16(2): 69. doi: 10.3390/md16020069
|
| [11] |
Coffey B M, Anderson G G. 2014. Biofilm formation in the 96-well microtiter plate. In: Filloux A, Ramos JL, eds. Pseudomonas Methods and Protocols. New York: Humana Press, 631–641, doi: 10.1007/978-1-4939-0473-0_48
|
| [12] |
Costerton J W, Lewandowski Z, Caldwell D E, et al. 1995. Microbial biofilms. Annual Review of Microbiology, 49: 711–745. doi: 10.1146/annurev.mi.49.100195.003431
|
| [13] |
De Siqueira Melo R, Brasil S L D C, De Carvalho L J, et al. 2016. Assessment of the antifouling effect of exopolysaccharides incorporated into copper oxide-based organic paint. International Journal of Electrochemical Science, 11(9): 7750–7763
|
| [14] |
Decho A W. 1990. Microbial exopolymer secretions in ocean environments: their role(s) in food webs and marine processes. Oceanography and Marine Biology, 28: 73–153
|
| [15] |
Delbarre-Ladrat C, Sinquin C, Lebellenger L, et al. 2014. Exopolysaccharides produced by marine bacteria and their applications as glycosaminoglycan-like molecules. Frontiers in Chemistry, 2: 85. doi: 10.3389/fchem.2014.00085
|
| [16] |
Dogan N M, Doganli G A, Dogan G, et al. 2015. Characterization of extracellular polysaccharides (EPS) produced by thermal Bacillus and determination of environmental conditions affecting exopolysaccharide production. International Journal of Environmental Research, 9(3): 1107–1116. doi: 10.22059/IJER.2015.998
|
| [17] |
Dubois M, Gilles K, Hamilton J K, et al. 1951. A colorimetric method for the determination of sugars. Nature, 168(4265): 167. doi: 10.1038/168167a0
|
| [18] |
Elbendary A A, Hessain A M, El-Hariri M D, et al. 2018. Isolation of antimicrobial producing Actinobacteria from soil samples. Saudi Journal of Biological Sciences, 25(1): 44–46. doi: 10.1016/j.sjbs.2017.05.003
|
| [19] |
Flemming H C, Neu T R, Wozniak D J. 2007. The EPS matrix: the “house of biofilm cells”. Journal of Bacteriology, 189(22): 7945–7947. doi: 10.1128/JB.00858-07
|
| [20] |
Gonzalez-Gil G, Thomas L, Emwas A H, et al. 2015. NMR and MALDI-TOF MS based characterization of exopolysaccharides in anaerobic microbial aggregates from full-scale reactors. Scientific Reports, 5: 14316. doi: 10.1038/srep14316
|
| [21] |
Gu Di, Jiao Yingchun, Wu Jianan, et al. 2017. Optimization of EPS production and characterization by a halophilic bacterium, Kocuria rosea ZJUQH from Chaka Salt Lake with response surface methodology. Molecules, 22(5): 814. doi: 10.3390/molecules22050814
|
| [22] |
He Jinzhe, Zhang Anqiang, Ru Qiaomei, et al. 2014. Structural characterization of a water-soluble polysaccharide from the fruiting bodies of Agaricus bisporus. International Journal of Molecular Sciences, 15(1): 787–797. doi: 10.3390/ijms15010787
|
| [23] |
Hwang G, Kang S, El-Din M G, et al. 2012. Impact of an extracellular polymeric substance (EPS) precoating on the initial adhesion of Burkholderia cepacia and Pseudomonas aeruginosa. Biofouling, 28(6): 525–538. doi: 10.1080/08927014.2012.694138
|
| [24] |
Jiang Peng, Li Jingbao, Han Feng, et al. 2011. Antibiofilm activity of an exopolysaccharide from marine bacterium Vibrio sp. QY101. PLoS One, 6(4): e18514. doi: 10.1371/journal.pone.0018514
|
| [25] |
Jiao Yongqin, Cody G D, Harding A K, et al. 2010. Characterization of extracellular polymeric substances from acidophilic microbial biofilms. Applied and Environmental Microbiology, 76(9): 2916–2922. doi: 10.1128/AEM.02289-09
|
| [26] |
Kanamarlapudi S L R K, Muddada S. 2017. Characterization of exopolysaccharide produced by Streptococcus thermophilus CC30. BioMed Research International, 2017: 4201809. doi: 10.1155/2017/4201809
|
| [27] |
Kavita K, Mishra A, Jha B. 2011. Isolation and physico-chemical characterisation of extracellular polymeric substances produced by the marine bacterium Vibrio parahaemolyticus. Biofouling, 27(3): 309–317. doi: 10.1080/08927014.2011.562605
|
| [28] |
Kavita K, Singh V K, Mishra A, et al. 2014. Characterisation and anti-biofilm activity of extracellular polymeric substances from Oceanobacillus iheyensis. Carbohydrate Polymers, 101: 29–35. doi: 10.1016/j.carbpol.2013.08.099
|
| [29] |
Lee M E, Lee H D, Suh H H. 2015. Production and characterization of extracellular polysaccharide produced by Pseudomonas sp. GP32. Journal of Life Science, 25(9): 1027–1035. doi: 10.5352/JLS.2015.25.9.1027
|
| [30] |
Lembre P, Lorentz C, Di Martino P. 2012. Exopolysaccharides of the biofilm matrix: a complex biophysical world. In: Karunaratne D N, ed. The Complex World of Polysaccharides. Croatia: IntechOpen, 371–392, doi: 10.5772/51213
|
| [31] |
Liu F C, Su C R, Wu T Y, et al. 2011. Efficient 1H-NMR quantitation and investigation of N-acetyl-D-glucosamine (GlcNAc) and N,N′-diacetylchitobiose (GlcNAc)2 from chitin. International Journal of Molecular Sciences, 12(9): 5828–5843. doi: 10.3390/ijms12095828
|
| [32] |
Mallick I, Bhattacharyya C, Mukherji S, et al. 2018. Effective rhizoinoculation and biofilm formation by arsenic immobilizing halophilic plant growth promoting bacteria (PGPB) isolated from mangrove rhizosphere: a step towards arsenic rhizoremediation. Science of the Total Environment, 610–611: 1239–1250. doi: 10.1016/j.scitotenv.2017.07.234
|
| [33] |
Maréchal J P, Hellio C. 2009. Challenges for the development of new non-toxic antifouling solutions. International Journal of Molecular Sciences, 10(11): 4623–4637. doi: 10.3390/ijms10114623
|
| [34] |
More T T, Yadav J S S, Yan S, et al. 2014. Extracellular polymeric substances of bacteria and their potential environmental applications. Journal of Environmental Management, 144: 1–25. doi: 10.1016/j.jenvman.2014.05.010
|
| [35] |
Palomo S, González I, De La Cruz M, et al. 2013. Sponge-derived Kocuria and Micrococcus spp. as sources of the new thiazolyl peptide antibiotic kocurin. Marine Drugs, 11(4): 1071–1086. doi: 10.3390/md11041071
|
| [36] |
Powell L C, Pritchard M F, Ferguson E L, et al. 2018. Targeted disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate oligosaccharides. NPJ Biofilms and Microbiomes, 4: 13. doi: 10.1038/s41522-018-0056-3
|
| [37] |
Pradeepa, Shetty A D, Matthews K, et al. 2016. Multidrug resistant pathogenic bacterial biofilm inhibition by Lactobacillus plantarum exopolysaccharide. Bioactive Carbohydrates and Dietary Fibre, 8(1): 7–14. doi: 10.1016/j.bcdf.2016.06.002
|
| [38] |
Qian Peiyuan, Chen Lianguo, Xu Ying. 2013. Mini-review: molecular mechanisms of antifouling compounds. Biofouling, 29(4): 381–400. doi: 10.1080/08927014.2013.776546
|
| [39] |
Rajasree V, Sunjaiy Shankar C V, Satheesh S, et al. 2014. Biofilm inhibitory activity of extracellular polymeric substance produced by Exiguobacterium sp. associated with the polychaete Platynereis dumerilii. Thalassas, 30(2): 13–19
|
| [40] |
Rani R P, Anandharaj M, Sabhapathy P, et al. 2017. Physiochemical and biological characterization of novel exopolysaccharide produced by Bacillus tequilensis FR9 isolated from chicken. International Journal of Biological Macromolecules, 96: 1–10. doi: 10.1016/j.ijbiomac.2016.11.122
|
| [41] |
Ricou P, Pinel E, Juhasz N. 2005. Temperature experiments for improved accuracy in the calculation of polyamide-11 crystallinity by X-ray diffraction. In: International Centre for Diffraction Data, ed. Advances in X-ray Analysis, Vol 48. Newtown, Pennsylvania, USA: International Centre for Diffraction Data, 171–175
|
| [42] |
Salama A J, Satheesh S, Balqadi A A. 2018. Antifouling activities of methanolic extracts of three macroalgal species from the Red Sea. Journal of Applied Phycology, 30(3): 1943–1953. doi: 10.1007/s10811-017-1345-6
|
| [43] |
Satheesh S, Ba-akdah M A, Al-Sofyani A A. 2016. Natural antifouling compound production by microbes associated with marine macroorganisms—A review. Electronic Journal of Biotechnology, 21: 26–35. doi: 10.1016/j.ejbt.2016.02.002
|
| [44] |
Satheesh S, Soniamby A R, Shankar C V S, et al. 2012. Antifouling activities of marine bacteria associated with sponge (Sigmadocia sp.). Journal of Ocean University of China, 11(3): 354–360. doi: 10.1007/s11802-012-1927-5
|
| [45] |
Sayem S A, Manzo E, Ciavatta L, et al. 2011. Anti-biofilm activity of an exopolysaccharide from a sponge-associated strain of Bacillus licheniformis. Microbial Cell Factories, 10: 74. doi: 10.1186/1475-2859-10-74
|
| [46] |
Schultz M P, Bendick J A, Holm E R, et al. 2011. Economic impact of biofouling on a naval surface ship. Biofouling, 27(1): 87–98. doi: 10.1080/08927014.2010.542809
|
| [47] |
Shiyamala D S, Priya P, Sahadevan R. 2014. Pyrrolo [1, 2-A] Pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl)- and phenol, 2,4-Bis (1,1-dimethy ethyl) novel antibacterial metabolites from a marine Kocuria sp. SRS88: optimization and its application in medical cotton gauze cloth against bacterial wound pathogens. International Journal of Pharmaceutical Research and Development, 6(2): 44–55
|
| [48] |
Solmaz K B, Ozcan Y, Mercan Dogan N, et al. 2018. Characterization and production of extracellular polysaccharides (EPS) by Bacillus pseudomycoides U10. Environments, 5(6): 63. doi: 10.3390/environments5060063
|
| [49] |
Sutherland I W. 2001. Biofilm exopolysaccharides: a strong and sticky framework. Microbiology, 147(1): 3–9. doi: 10.1099/00221287-147-1-3
|
| [50] |
Thomas K V, Brooks S. 2010. The environmental fate and effects of antifouling paint biocides. Biofouling, 26(1): 73–88. doi: 10.1080/08927010903216564
|
| [51] |
Tian Yu. 2008. Behaviour of bacterial extracellular polymeric substances from activated sludge: a review. International Journal of Environment and Pollution, 32(1): 78–89. doi: 10.1504/IJEP.2008.016900
|
| [52] |
Viju N, Satheesh S, Punitha S M J. 2016. Antibiofilm and antifouling activities of extracellular polymeric substances isolated from the bacteria associated with marine gastropod Turbo sp. Oceanological and Hydrobiological Studies, 45(1): 11–19. doi: 10.1515/ohs-2016-0002
|
| [53] |
Vliegenthart J F G, van Halbeek H, Dorland L. 1981. The applicability of 500-mhz high-resolution 1H-NMR spectroscopy for the structure determination of carbohydrates derived from glycoproteins. Pure and Applied Chemistry, 53(1): 45–77. doi: 10.1351/pac198153010045
|
| [54] |
Wang Chunlei, Fan Qiuping, Zhang Xiaofei, et al. 2018. Isolation, characterization, and pharmaceutical applications of an exopolysaccharide from Aerococcus Uriaeequi. Marine Drugs, 16(9): 337. doi: 10.3390/md16090337
|
| [55] |
Wang Kailing, Wu Zehong, Wang Yu, et al. 2017. Mini-review: antifouling natural products from marine microorganisms and their synthetic analogs. Marine Drugs, 15(9): 266. doi: 10.3390/md15090266
|
| [56] |
Wang Ji, Zhao Xiao, Tian Zheng, et al. 2015. Characterization of an exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibet Kefir. Carbohydrate Polymers, 125: 16–25. doi: 10.1016/j.carbpol.2015.03.003
|
| [57] |
Wu Shimei, Liu Geihua, Jin Wengyuan, et al. 2016. Antibiofilm and anti-infection of a marine bacterial exopolysaccharide against Pseudomonas aeruginosa. Frontiers in Microbiology, 7: 102. doi: 10.3389/fmicb.2016.00102
|
| [58] |
Yadav V, Prappulla S G, Jha A, et al. 2011. A novel exopolysaccharide from probiotic Lactobacillus fermentum CFR 2195: Production, purification and characterization. Biotechnology and Bioengineering, 1(4): 415–421
|
| [59] |
Yang L H, Miao Li, Lee O O, et al. 2007. Effect of culture conditions on antifouling compound production of a sponge-associated fungus. Applied Microbiology and Biotechnology, 74(6): 1221–1231. doi: 10.1007/s00253-006-0780-0
|
| [60] |
Yildiz H, Karatas N. 2018. Microbial exopolysaccharides: resources and bioactive properties. Process Biochemistry, 72: 41–46. doi: 10.1016/j.procbio.2018.06.009
|
| [61] |
Zhang Li, Liu Chunhong, Li Da, et al. 2013. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. International Journal of Biological Macromolecules, 54: 270–275. doi: 10.1016/j.ijbiomac.2012.12.037
|

Figures(7) / Tables(2)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: